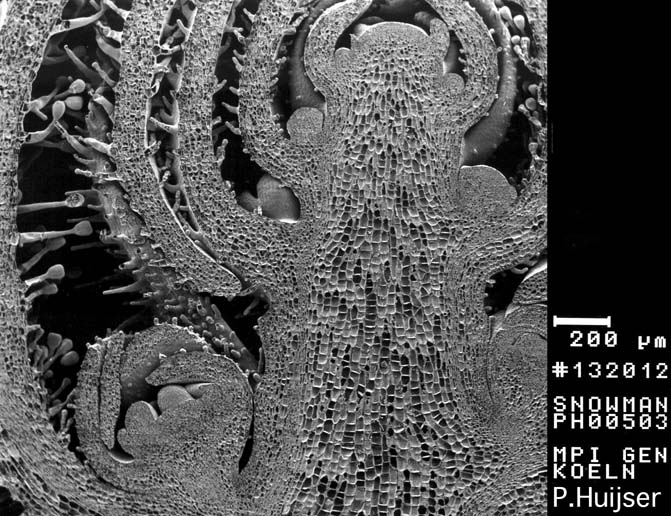
Scanning Electron Micropgraph: Longitudinal section through an inflorescence apex - Snapdragon (Antirrhinum majus))
Botany online 1996-2004. No further update, only historical document of botanical science!
revised by Martina
HÖFT, Nairobi
Cormophytes produce new cells throughout their life and form new organs at regular intervals (some tallophytes do the same). All cells of an embryo display the same activity of division, but as soon as a certain state of differentiation and thus a certain size has been reached, the production of new cells is restricted to special tissues, the meristems.
Typical meristems are found at the tips of all stems and roots. They are called apical meristems. Those of the shoot are usually protected by involucral leaves and the whole complex forms a bud.
Scanning Electron Micropgraph: Longitudinal section through an inflorescence apex - Snapdragon (Antirrhinum majus))
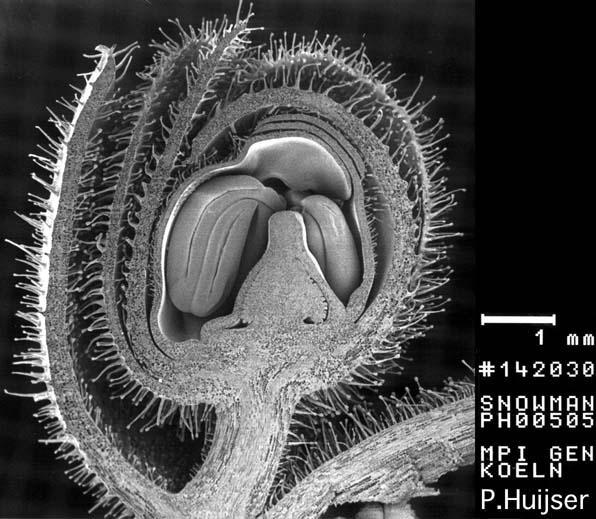
Scanning Electron Micrograph: Immature Flower Bud - Snapdragon (Antirrhinum majus)
The apical meristems are the cause of tip growth that is one of the most striking features of vegetative growth. This contrasts with animal growth, that is an allometric process, i.e. all parts of the body grow in a weay that maintains the body's overall proportions. As soon as an animal has reached its final size, growth ceases. This, however, does not mean that no more cell divisions are taking place. Cell divisions occur all the time, especially in epithelia and in blood stem cells. But this division activity has always the function of replacing old or damaged cells. No such repair occurs in plants. If a leaf or flower or any other part is damaged, it is neither replaced nor repaired. Instead, new organs grow, and this again does not happen in animals.
The cells of a meristem have a potentially unlimited ability to divide. But division is controlled. Many plants, for example, follow the principle of apical dominance, where the activity of the lateral meristems is suppressed while the tip is growing. In this case, information is exchanged between the two tissues in question. If the tip of the main axis is cut off, suppression is abolished and the lateral buds start to sprout.
Many plants like gymnosperms and dicots have extensive lateral meristems (the cambium, the vascular cambium, and the cork cambium) that give rise to growth in girth, also called secondary growth. Their activity may fluctuate in the course of the year and is reflected by annual rings.
Other plants, like monocots, have intercalary meristems. They are actively growing meristems that differ clearly from apical meristems and are located between more or less differentiated tissues usually at the base of each internode. The formation of secondary meristems shows that differentiated cells can reverse their state and go back to a meristematic existence. Their ability to divide is thus not lost, though it is not used unless an adjustment to changed circumstances requires it. The ability of indeterminate growth is widely exploited by the use of cuttings in horticulture. It is also displayed in the formation of roots at the cutting sites of begonia leaves, for example.
But what exactly is a meristem ? The following three examples will help to answer this question.
The marine brown alga Dictyota dichotoma consists of a flat thallus with a thickness of three cell layers. The large cell at its apical tip divides periclinal, while the subapical progenitor cells divide in an anticlinal manner. The progenitor cells themselves display alternating peri- and anticlinal cell division. The thallus enlarges thus both in length and in width. At regular intervalls, the apical cell divides in an anticlinal manner, too, resulting in two apical cells and a subsequent forking of the thallus. Although this example explains the way a division scheme works, it does not explain where the meristematic cells end and where the differentiated ones begin. The classical scheme of a cormophyte given by SACHS says that no clear demarcation exists. Instead, there is a gradient between meristematic and differentiated cells. The meristematic properties are thus not lost during one division, but they decline gradually.
The second example shows how division takes place in some liverworts, in simple cryptogams like horsetail and in many ferns. Contrary to that of many algae, their cormus normally resembles a three-dimensional body with a single tetraedical initial cell at the tip of the axis. It produces progenitor cells by regularly alternating the planes of division. These progenitor cells are given off towards the base. The progenitor cells may again divide in both anti- and periclinal manners.
The tip of phanerogams, my third example, is formed by a whole group of cells. They are not specialized and therefore called vegetation zone (see illustrations of Acacia, Oxypolis, Wheat). The apical meristem is organized into several cell layers. In angiosperms and in some gymnosperms, it has to be distinguished between the outer tunica and the central corpus (A. SCHMIDT, 1924; F. A. L. CLOWES, 1924; A. FÖRSTER, 1943). The meristematic cells divide mostly in an anticlinal manner, while those of the corpus display both planes of division: anti- and periclinal. Some parts of the corpus are dominated by a certain plane of division, thereby determining the first step of differentiation. Usually, the outer layer of the tunica produces the epidermis. The inner tissues of the plant stem from the corpus, the tunica or both.
These schemes are useful guidelines for the understanding of the plant's morphogenesis, since they point out the importance of the information that is contained in the cell's position within the tissue. A cell will only develop into a certain direction, if its position within the tissue is "right". If a cell is transferred to a new position, it will adopt new functions and thus dedifferentiate. At the beginning of the 20th century, G. HABERLAND (Universität Graz, later Berlin) put forward the sentence of the totipotency of plant cells. It states, that each cell of a plant keeps the ability to develop a complete plant. This is true for most, but not all plant cells. And still: growth through division is in fact characteristic for the meristematic state of a cell, but it is by no means restricted to it. A meristem does not only include the initial cells and their immediate progenitors, but also some parts of the shoot.
The situation is similar in root meristems, although their anatomy is different. The shoot apex is protected by the involucral leaves. In roots, this function is exerted by the root cap. Its cells are produced by a specialized root cap meristem and are progressively replaced towards the tip. These cells have to have a high turn-over, since the outmost cell layer is easily damaged by soil particles as a result of the growth movement of the root.
It is normally distinguished between primary and secondary growth of the shoot. The first is the phase of plant development that gives rise to new organs and to the basic plant form, the latter brings about growth in girth and the formation of new vascular tissues (A. de BARY, 1877).
The procambium is the meristematic tissue that produces the primary vascular tissues: xylem and phloem. It develops directly beneath the growing tip next to new leaf primordia. The development of new leaves is thus tightly connected with that of new vascular tissues. The vascular tissues of leaves are also called leaf veins. The cells of the procambium are generally combined in cords. They form an extension of the vascular tissues into the growing tip and do thus provide the connection of the newly developed organ with the conductive systems of the plant. The cells of the procambium are elongated and become even more so in the course of their development. The volume of their vacuoles increases considerably, lending them a lighter and more transparent appearance than their neighboring cells. This is decisive for their differentiation into xylem or phloem cells.
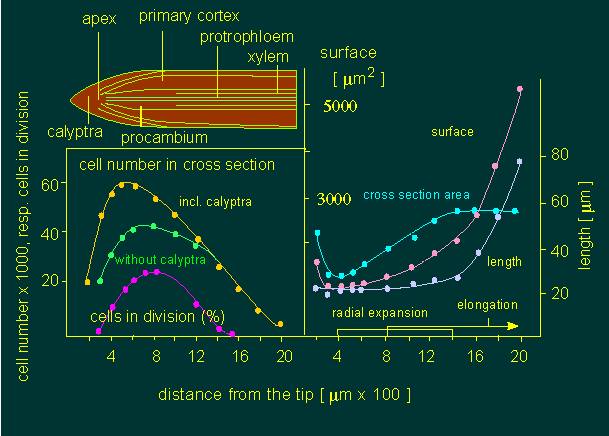
Growth processes within the first two millimetres of a radical apex of Allium cepa (onion). The following data was collected to characterize the growth occurring at the given stages. On the left: number of cells within a 100 µm thick cross-section. On the right: area of cross-section; length and surface of the cells at different distances from the radical apex (according to W. A. JENSEN and M. ASHTON, 1960).
Some of the meristematic cells in plants with secondary growth keep their meristematic state and become cells of the cambium.
What are the causes for the development of the procambium? Two models exist:
- They are to be found in the cell-cell interactions that take place between the cells of the bud, that determine the pattern of the leaf positions and the positions of xylem and phloem.
- Induction is achieved by fully differentiated tissues. The necessary information is conducted to the bud via already differentiated cells of the procambium.
The first model was confirmed experimentally, since formation and further development of the procambium are undisturbed in isolated bud meristems. The determination of cells takes place before any morphological change can be seen. Differentiation describes the (usually irreversible) changes of structures and functions that result in a certain specialization, while determination means the (usually irreversible) triggering of processes that lead to these changes.
The cambium is the prototype of a lateral meristem. It is mono- or multilayered depending on its origin and forms normally a continuous cell layer of tubular shape that is located at the periphery of the shoot or root. It separates xylem from phloem, if present. The cambium develops from the procambium next to the vessels, ensuring a continuity of the meristematic state.
The cambium between the vessels derived from already differentiated, parenchymatous cells. The first type is also called fascicular (within the vascular bundle) cambium, while the latter one is termed interfascicular (between the vascular bundles). The interfascicular cambium is a secondary meristem. A few monocots (Dracaena, Yucca, Aloe and others) have again another type of cambium, the extrafascicular cambium.
The cells of the cambium are often termed initials, since they initiate the formation of specialized progenitor cells after division. Two cell types occur within the cambium:
- Fusiform initials. They are the mother cells of all xylem and phloem elements, as well as of all other cells that are oriented parallel to the organ's axis. They are flat, elongated with pointed ends and highly vacuolate. It is their spindle-like shape that caused their name.
- Ray initials are nearly isodiametric, small cells that occur often in groups. They develop from fusiform initials or their progenitors. They produce the radially orientated rays in wooden plants (transversal elements).
I. W. BAILEY, who did the basic work on the organization of the cambium in the 20th of this century, gained the following data when comparing a one year old and a sixty years old stem of Pinus strobus (Weymouths Pine).
|
|
||
|
|
|
|
|
|
|
|
|
radius of the tube |
|
|
|
girth of the cambium |
|
|
|
average length of fusiform initials |
|
|
|
number of fusiform initials in |
|
|
|
number of ray initials |
|
|
|
|
||
His data show that both length and number of fusiform initials increases with the age of the stem. The increase in number and the widening (dilatation) of the cambium tube is caused by the widening of the central xylem cylinder, that is produced by the cambium itself.
Secondary vascular elements are given off into opposite directions by periclinal (tangential) division of fusiform cells. The developing xylem elements grow towards the inner cylinder, that of the phloem towards the outer of the cylinder. The progenitors of the initials are organized in radial rows that make it easy to trace back their descent. The increase in girth is compensated by the anticlinal divisions occurring at regular intervals. The activity of the cambium is temperature-dependent: it causes the annual rings. Their thickness, i.e. the activity of the cambium, is determined by extern factors like temperature, day length, soil humidity, temperature and others. Besides its function in the production of vascular tissues, the cambium has an additional capacity for the healing of wounds.
The cork cambium (or phellogen) is a secondary lateral meristem that serves to produce the secondary outer surface, the bark, that replaces the epidermis. It is without exception given off towards the outward direction. Often, though not always, the cork cambium produces cells towards the inner of the stem, that form the phelloderm.
The cork cambium has a rather simple structure, if compared to the cambium. In cross-section, the cells have a right-angled shape, they are flat in radial and tangential sections. Their plasma is highly vacuolated and may contain chloroplasts and tannic acids.
The cork cambium stems from epidermal cells or/and from cells of the underlying parenchyma. It is distinguished between primary and secondary cork cambium taking into account the fact that it can be assembled within the stem several times during a plant's life. The cork cambium is monolayered during the first year in some species and becomes multilayered later on. It may be active for several years, sometimes even for the whole life or for just one year. Its activity is, just as that of the cambium, influenced by extern factors.