Botany online 1996-2004. No further update, only historical document of botanical science!
Phototropism is a growth movement induced by a light stimulus. Growth towards a source of light is called positive phototropism, that away from the source is termed negative phototropism. The tips of shoots are usually positively, that of roots negatively phototropic.
Originally (with J. v. SACHS, for example) was phototropism called heliotropism, because the plant grows towards the sun. The name was altered when it became clear that plants react also towards artificial sources of light (W. PFEFFER). Before proceeding further shall another problem be outlined shortly: Does the plant react to light or to the air? Here is a citation from A. de CANDOLLE ( 1834/38).
"Gardeners and farmers tell usually that plants are attracted by the air and only TESSIER proved this to be false by a simple experiment: He placed living plants in a basement with two openings, on one side spent a glass window light but no air, on the other brought an airhole leading through a spacious and dark room air but not light. All plants grew towards the glass window."
A.de CANDOLLE noticed as soon as 1809 that the growth towards light is caused by an unequal growth of the opposite parts of an organ. The part exposed to light grows slower than the one that is not exposed. J. v. SACHS discovered the importance of the light quality (the dependence on the wave length) for the phototropic reaction. Blue, violet and ultraviolet light together have the same effect as very strong white light. The effect remains even after the UV part is taken away. Red, yellow , and green light has no effect on most plants. Red light causes a phototropic reaction in some fern prothalliums, though.
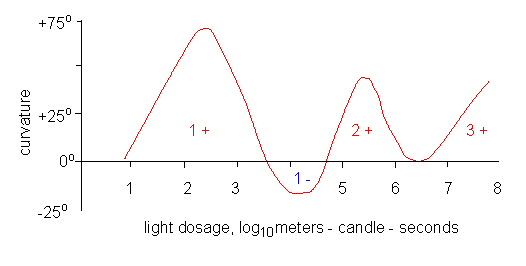
The amount of light and the phototropic reaction are linked. This connection is known in literature under the terms Bunsen-Roscoe law, product law or reciprocity theorem (FRÖSCHEL and BLAAUW, 1908, BLAAUW, 1909). The Bunsen-Roscoe law states that the product of time and intensity, and thus the energy amount of the used light, is the measure of the stimulus strength. It is thus of no importance whether a light stimulus of low intensity is applied for a longer period of time or whether a stimulus of high intensity is applied for a short time. We know today that the Bunsen-Roscoe law applies only in a very limited sense, since a certain minimal amount of light has to be present in order to trigger the reaction (threshold value), and an increase in light intensity does not always cause an increase in the phototropic reaction, but may in contrast suppress the positive reaction. A continuous increase causes a second positive reaction with a new maximum, that decreases again and rises once more (= 1., 2., and 3. positive phototropic bending).
The dose-effect curve of the phototropism of etiolated Avena coleoptiles (schematic depiction). The amount of white light is given in lux seconds, the ordinate gives the positive or negative bending. The reaction spheres of the 1., 2., and 3. positive, as well as that of the negative phototropic bending are shown (according to H. MOHR and P. SCHOPFER, 1978, and H. G. duBUY and E. NUERNBERGK, 1943).
Positive and negative phototropism within the same tissue are characteristic not only for Avena coleoptiles. It can also be observed under natural light conditions with germinating plants of the tropic Aracea Monstera gigantea. At low amounts of light (a low flow of photons) does it react positively, at too high amounts negatively phototropic. It differs from Avena primarily in the stimulus thresholds at which one reaction becomes the opposite.
Phototropic reactions are characteristic for growing tissues, and are less easily detected in fully differentiated ones. This is on one hand caused by the cells’ loss of plasticity, and on the other hand by the development of mostly inflexible strengthening elements that set a mechanic resistance against each deformation of the tissue. In the shoot axis are they arranged in the periphery thus bringing about an especially high stability.
These facts show that we have stimulus perception, stimulus forwarding, the mechanism of the bending, and experimental data for all three reaction parts, but it is still not understood how these parts are linked. We do not know how the synthesis of the information molecules (the phytohormone auxine) is influenced by the light receptor after stimulation. We do not know either how the decision for a negative or a positive phototropic reaction is made, and we have only rudimentary knowledge about the way in which auxine could stimulate the elongation of the cell walls in the growth zone.
A further example of literature is the phototropic bending of the sporangium of Phycomyces: Indeed is its mechanism better understood than that of the Avena coleoptile, since the problem of intracellular forwarding of information does not occur. The question how opposite walls of the same cell can grow with differing velocity remains. Changes of the turgor cannot explain it.