Botany online 1996-2004. No further update, only historical document of botanical science!
The third Mendelian law states that hereditary factors are inherited independently. This statement holds true for all seven features analyzed by MENDEL. His correspondence with C. v. NÄGELI nevertheless mentions briefly the fact that this law is sometimes broken. Such an example was found by W. BATESON, E. R. SAUNDERS and R. C. PUNNETT, when they studied the sweet pea Lathyrus odoratus in 1905. They examined the inheritance of the flower colour and the pollen structure. 381 F2-plants were analyzed. The respective characters segregated as follows:
284 (= 74.6%) with crimson flowers and long pollen,
21 (= 5.5%) with crimson flowers and round pollen,
21 (= 5.5%) with red flowers and long pollen and,
55 (= 14%) with red flowers and round pollen.
In a repeat of the experiment found R. C. PUNNETT in 1917 among 6952 F2 plants
4831 (= 69.5%) with crimson flowers and long pollen,
390 (= 5.6%) with crimson flowers and round pollen,
393 (= 5.6%) with red flowers and long pollen and
1338 (= 19.3%) with red flowers and round pollen.
This meant that the hereditary factors were almost always passed on in units. They are generally not separated during the production of the germ cells, so that the combinations:
flower colour: crimson,
pollen: long and
flower colour: red
and pollen: round
occur roughly in the ratio 3:1. The exceptions are conspicuous, because the coupling is sometimes broken and low percentages of reciprocal combinations (flower colour: crimson, pollen: round) and, in to the same amount, flower colour: red and pollen: long occur. The experiments gave no conclusive explanation for these findings. But in view of the chromosome theory, a plausible explanation can be given.
Later studies performed by T. H. MORGAN and his collaborators on the fruit fly Drosophila melanogaster showed that the number of coupling groups corresponds to the number of chromosomes in a haploid set and that occasional exceptions were caused by the exchange of chromosomal pieces that occurred during the pairing of the homologous chromosomes in meiosis. The results of these experiments became an important pillar of the chromosome theory and offered a way to localize genes on chromosomes by the analysis of their exchange frequency.
Some few years after the rediscovery of MENDEL's laws, C. CORRENS (1909) and E. BAUR (1909, 1910) described crossings, for which the Mendelian laws did not apply. A certain type of variegated leaves, for example, the green-white one of Mirabilis jalapa and Antirrhinum majus is inherited according to the following scheme:
This result breaks with the first Mendelian law. No segregation occurs in the F2 generation. This does also mean that MENDEL's second law does not apply.
These phenotypes depend on the maternal genotypes. In contrast to
the nucleus of the pollen, that of the egg is surrounded by a
voluminous plasma. The thought that it is this plasma that contains
the hereditary material, that influences the phenotypes in the
observed way, was thus not far-fetched. This type of inheritance was
therefore termed cytoplasmic or maternal, later also
extrachromosomal inheritance.
E. BAUR found a variation of this type of inheritance in Pelargonium zonale. This species has, besides plants with completely green leaves also such that have leaves with white or yellow margins. The inheritance of the yellow-rimmed type follows the same pattern as that of the plants with white rims. The cotyledons can already in the F1 generation be analyzed. In addition to green and spotted leaves, completely white plants do occur that cannot develop further due to the lack of chlorophyll. The results of the crossings are:
F1: 159 plants (of three independent experiments) were analyzed:
F1: 65 individuals were counted:
Although this example resembles that of Mirabilis jalapa
and Antirrhinum majus, it does moreover show that also the
(small amount of) plasma of the pollen has an influence on the
expression of features.
But what are plasmatic factors? Which conditions have to be fulfilled by the hereditary material? Certainly two very general criteria:
![]() They have to pass on the information contained in their
structure
They have to pass on the information contained in their
structure
![]() They have to be capable of replication. Lost material
cannot be replaced.
They have to be capable of replication. Lost material
cannot be replaced.

Evidence for Cytoplasmic Inheritance. The phenotype does not depend on the nuclear genome.
Not only the chromosomes, but also the DNA of the plastids meet these criteria. The morphological continuity of the plastids was proven by F. SCHMITZ, A. F. SCHIMPER and A. MEYER from 1882-1885. Based on their work, A. BAUR interpreted his results in 1909:
"The fertilized egg that has developed from the fusion of a 'green' with a 'white' germ cell contains thus two chromatophores (note: plastids), green and white ones. When the cell divisions of the zygote that has developed into the embryo take place, the chromatophores are dispersed onto the daughter cells according to the laws of probability. If a daughter cell inherits only white chromatophores, then all its descendants will have white chromatophores and thus produce a white piece of mosaic. If a daughter cell contains only green chromatophores, then a constant, green cell complex will develop. Cells with both types of chromatophores will continue to be able to segregate, etc. ...."
This conclusion led to the postulation of the segregation
hypothesis (E. BAUR, 1909, O. RENNER, 1922). It assumes that
different types of plastids live in one and the same cell at the same
time. That this is actually true was proven years later by the
observation of so-called 'mixed cells'.
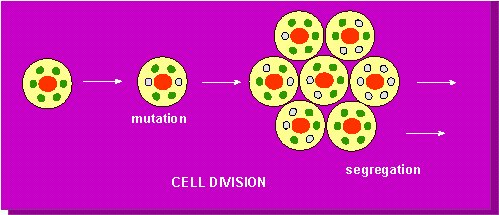
P. MICHAELIS of the Max-Plank-Institut für Züchtungsforschung, Köln-Vogelsang calculated in 1955/56 the statistical probability of the segregation of two different types of plastids under different conditions. Roughly 10-100 plastids and 700 mitochondria occur in the cells of higher plants. Under the assumption that the complete development of an annual plant takes about 100 consecutive cell divisions, a nearly complete segregation of the plastids can be predicted. This statements does not apply to mitochondria. Their segregation stays incomplete, since there are so many of them in one cell.
Mitochondria have to be mentioned explicitly, since they were also recognized as containers of genetic material. These results were mainly gained by studies on yeasts, other fungi, protozoa and animal cells. During the 1960s, it could finally be verified that both plastids and mitochondria contain DNA that itself contains genetic information.
Extrachromosomal inheritance can thus be characterized by three properties:
![]() The distribution does not follow Mendelian laws,
The distribution does not follow Mendelian laws,
![]() reciprocal crossings behave differently,
reciprocal crossings behave differently,
![]() the genetic material segregates during ontogenesis.
the genetic material segregates during ontogenesis.
But the white-green patches of leaves or variegated flowers are not always caused by the segregation of plasmatic factors, because
white-green colourings are in many cases caused by viruses; the same is true for some many-coloured flowers, like tulips. The patches can only be kept by vegetative propagation or grafting, since viruses do nearly never infect the seeds, so that the offspring is healthy (green leaves, all one coloured flowers).
A whole range of regularly structured green-white patterns follows Mendelian laws and is therefore determined by nuclear genes.
Contrary to the phenomenons just mentioned, patterns occur that look as if numerous changes would take place during ontogenesis. Green cells develop from a white cell line that again becomes green and so on. These phenomenons were formerly thought to be caused by so-called 'fragile' genes. Today is it known that there exist indeed 'jumping' genes that act like switches of other genes.
The existence of genes within the nucleus, the plastids (and the mitochondria) brings up the question of the co-operation between the respective genomes. Or, in other words: does every nucleus cooperate well with every plasma?
This is a situation that occurs always, when two different species cross. Sometimes, the pairing works and sometimes not. Often, the crossing of two different species results in a 'paleness of the hybrids' ("Bastardbleiche"), a phenomenon that was analyzed by O. RENNER during the 1920s and 1930s in species of the genera Oenothera. The reciprocal crossing of the two species Oenothera hookeri x Oenothera lamarckiana results in the following differences:
RENNER traced the differences back to differences in the genotypes of the plastids, that are influenced by the genotypes of the nuclei. Through extensive series of crossing experiments with a number of Oenothera-species, different types of plastids and different genome complexes were characterized during the last decades and the compatibilities of the respective combinations were tested (F. SCHÖTZ, Universität München, W. STUBBE, Universität Düsseldorf). The examination of nearly 400 combinations showed that the plastids of 14 wild species can be grouped into 5 reaction types (I - V). One of the differences is the speed of their propagation. The single Oenothera-species can thus be arranged according to their decreasing rate of propagation in the following row:
hookeri > bauri > lamarckiana > rubricaulis > suaveolens > biennis > syrticola > parviflora > rubricuspis > ammophila > artrovirens.
The speed of propagation is mainly determined by the plastid itself and not by the nucleus. The properties of the nuclei can also be classified (A-C). The results of the study were arranged in a scheme, which shows that a varying degree of compatibility between different nuclei and different plastids exists.
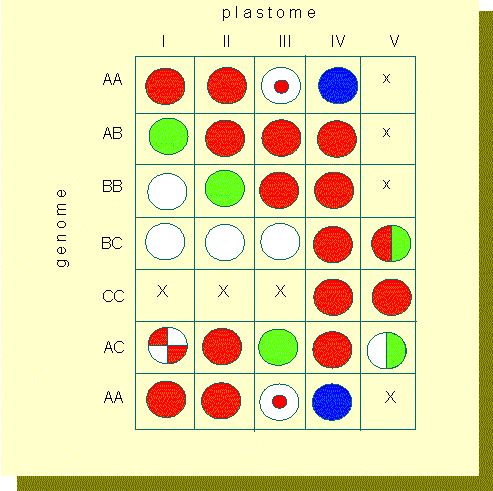
Combination square that offers a survey of the plastids ability to become green. Chlorophyll generation in species of the subgenera Euoenothera depends on the genome-plastid combinations. Red squares: full compatibility (green plastids), x: lethal combinations, white squares: combinations with non-green plastids, half squares: combinations with partly green plastids, squares with sectors or concentric rings: offspring of differing phenotypes (according to W. STUBBE, 1959).
The influence of a certain nuclear gene on the ability of the corn plastids to become green was studied in 1946 by M. M. RHOADES (Cold Spring Harbor Laboratory). The gene Iojap (Ij) causes (regular) green-white stripes on the corn leaves, when it is present in a homozygous condition. At first, it looked as if it would behave according to the Mendelian laws. But detailed crossing experiments lead to the following results:
F1: uniformly green (Ijij)
F1: mostly green (Ijij), in addition
striped (Ijij) and white (lethal) (Ijij)
Although the genotype is always the same does it result in different phenotypes.
green (IjIj)
striped (IjIj)
white (lethal) (IjIj)
green (Ijij)
striped (Ijij)
white (lethal) (Ijij).
The analysis of the results allows the conclusion that the gene ij
induced the development of abnormal chloroplasts that manifest
themselves independently of the constitution of the nucleus. That is
the cause for the occurrence of plants with the genotype IjIj or Ijij
that are either striped or white. But how do the stripes develop? Why
do not only defect plastids (white) develop in the presence of ijij?
Obviously, a number of plastids stay green also in the ijij-plants
and it looks, as if the colourless ones, too, could sometimes become
green. Anyway, the results show that the genetic information of the
nucleus and the plastids supplement each other and that the genomes
of the different compartments work together.
Molecular-genetic studies performed during the last years revealed that a perfectly working, exactly attuned flow of information between nucleus and plastids (and between nucleus and mitochondria) exists and that the genes of the chloroplasts and mitochondria exert a decisive and irreplaceable influence on the existence of the cells. Genes that are localized in chloroplasts run reactions of photosynthesis, those in the mitochondria the sequence of reactions of the respiratory chain.