Evolution of Eukaryotes | Eukaryotic Organelles and Prokaryotic Symbionts
Classification of Protists | Protozoa: Single-Celled, Motile Organisms | Algae
The transition to eukaryotic cells appears to have occurred during the Proterozoic Era, abbut 1.2 billion years ago. However, recent genetic studies suggest eukaryotes diverged from prokaryotes closer to 2 billion years ago. Fossils do not yet agree with this date.
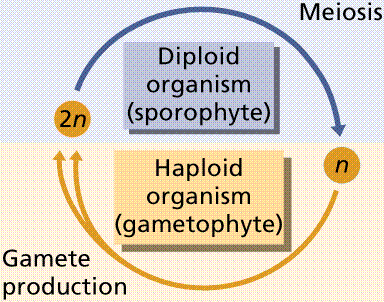
Protists exhibit a great deal of variation in their life histories (life cycles). They exhibit an alternation between diploid and haploid phases that is similar to the alternation of generations found in plants. Protist life cycles vary from diploid dominant, to haploid dominant.
Image from W.H. Freeman and Sinauer Associates, used by permission.
The great diversity of form, habitat, mode of nutrition, and life history exhibited by eukaryotes suggests they evolved several times from various groups of prokaryotes. This makes the Protista a polyphyletic group. Eukaryotes are generally larger, have a variety of membrane-bound organelles, greater internal complexity than prokaryotic cells, and has a secialized method of cell division (meiosis) that is a prelude to true sexual reproduction. Protists might be viewed as a group from which the other eukaryotic kingdoms evolved.
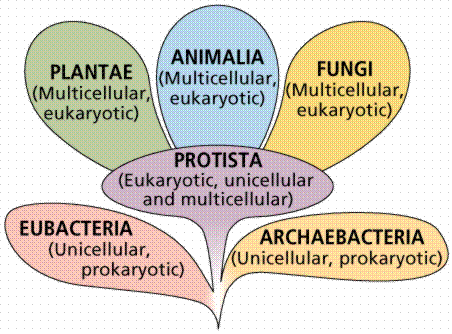
Image from W.H. Freeman and Sinauer Associates, used by permission.
Symbiosis is the interactive association of two or more species living together. There are several types of symbiosis in Nature. Parasitism is a symbiosis where one organism causes harm to the other, its host. Commensalism is a symbiosis where one organism benefits and the other is not harmed or helped. Mutualism is a symbiosis where both organisms benefit.
The symbiotic model proposed by Lynn Margulis suggests possible symbiosis of bacteria within early eukaryotic cells. Margulis proposed the mechanism of endosymbiosis to explain the origin of mitochondria and chloroplasts from permanent resident prokaryotes. According to this idea, a larger prokaryote (or perhaps early eukaryote) engulfed or surrounded a smaller prokaryote some 1.5 billion to 700 million years ago.
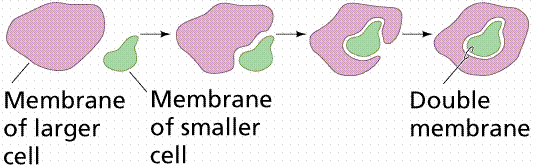
Image from W.H. Freeman and Sinauer Associates, used by permission.
Instead of digesting the smaller organisms the large one and the smaller one entered into a type of symbiosis known as mutualism, where both organisms benefit and neither is harmed. The larger organism gained excess ATP provided by the "protomitochondrion" and excess sugar provided by the "protochloroplast", while providing a stable environment and the raw materials the endosymbionts required. This is so strong that now eukaryotic cells cannot survive without mitochondria (likewise photosynthetic eukaryotes cannot survive without chloroplasts), and the endosymbionts cannot survive outside their hosts. Nearly all eukaryotes have mitochondria. Mitochondrial division is remarkably similar to the prokaryotic methods that will be studied later in this chapter. A summary of the theory is available by clicking here.
Chloroplasts and mitochondria still retain their own DNA, and may have evolved several times. The photosynthetic pigments (Table 1) in the red, brown, golden-brown, and green algae are very different, leanding support for the idea of several different endosymbiotic events.
Table 1. Photosynthetic pigments of algae and plants. Prokaryote groups are shown in red, protists in blue, and vascular plants in purple.
Cyanobacteria chlorophyll a, chlorphyll c, phycocyanin,
phycoerythrin Chloroxybacteria chlorophyll a, chlorphyll b Green Algae (Chlorophyta) chlorophyll a, chlorphyll b,
carotenoids Red Algae (Rhodophyta) chlorophyll a, phycocyanin, phycoerythrin,
phycobilins Brown Algae (Phaeophyta) chlorophyll a, chlorphyll c, fucoxanthin and other
carotenoids Golden-brown Algae (Chrysophyta) chlorophyll a, chlorphyll c, fucoxanthin and other
carotenoids Dinoflagellates (Pyrrhophyta) chlorophyll a, chlorphyll c, peridinin and
other carotenoids Vascular Plants chlorophyll a, chlorphyll b,
carotenoids
The DNA, ribosomes, biochemistry, and reproduction of chloroplasts and mitochondria are remarkably bacteria-like. Some living eukaryotes, such as the amoeba Pelomyxa, lack mitochondria, having instead endosymbiotic bacteria that perform "mitochondrial duties" of ATP generation.
The protists include heterotrophs, autotrophs, and some organisms that can vary their nutritional mode depending on environmental conditions. Protists occur in freshwater, saltwater, soil, and as symbionts within other organisms. Due to this tremendous diversity, classification of the Protista is difficult.
The first protist fossils occur in rocks approximately 1.2-1.4 billion years old from the Bitter Springs Formation in Australia. Multicellular protists appeared in the fossil record more than 600 million years ago. Some problemmatic fossils, thought by some paleobotanists to be algae, have been found in rocks approximately one billion years old.
This group of protists are single-celled, motile, heterotrophs. Most digest their food by vacuoles formed by phagocytosing other organisms (bacteria or other single-celled creatures). Reproduction varies greatly, from a binary fission-like process to true meiosis. The main distinguishing feature is the method of locomotion: flagella, cilia, or pseudopodia.
Amoeba and Pelomyxa move by extensions of their cytoplasm known as pseudopodia. Pseudopodia are used by many cells, and are not fixed structures like flagella but rather are associated with actin near the moving edge of the cytoplasm.
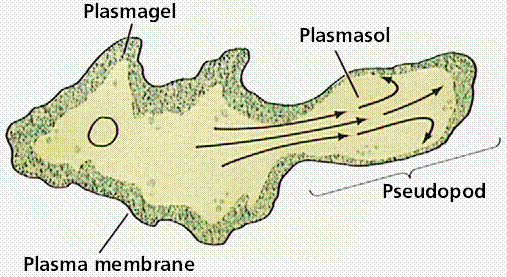
Image from W.H. Freeman and Sinauer Associates, used by permission.
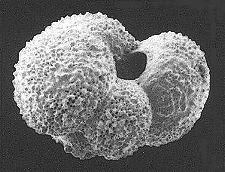
Foraminifera (forams) live in the oceans and secrete a shell (also known as a test) composed of silica or calcium carbonate. Thus, the fossil record of forams is quite good. Oxygen isotope data from forams has been used to calculate ocean temperature fluctuations over the past 100,000 years.
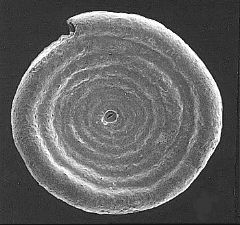
These images of recent forams are from http://www.cs.uwindsor.ca/meta-index/fossils/forams/gsls/ss-master-fsurface.html. Ammodiscus catinus (L), Globigerina bulloides (R).
Foraminiferan Test, Ephidium sp. (SEM x770). This image is copyright Dennis Kunkel http://www.pbrc.hawaii.edu/~kunkel/gallery, used with permission.
Entamoeba hystolitica is an intestinal parasite in humans that causes amoebic dysentery (also known as Montezuma's Revenge or the Aztec Two-step). It is present in the water supply of many communities in Mexico (and other countries), and unless specifically filtered, toxins from this amoeba will cause a disease that can ruin a vacation. Over time, your body will acclimate to the toxins, but since many of us only are exposed for short times, our bodies will not be able to cope. Drinking filtered water should prevent contacting this illness. The Bad Bug Book website has a description of this illness.
The flagellates use one or more flagella for locomotion. They typically lack cell walls and chloroplasts, being in general heterotrophic. Trypanosomes require two hosts, one a mammal, to complete their life cycle, and cause the diseases African sleeping sickness, Chagas disease, and leishmaniasis. Trichonymphs are symbionts inside the intestines of termites.
Members of this group cause malaria and toxoplasmosis. The life history of each organism has it infecting a different host for part of its growth. Toxoplasmosis is transmitted from cats to humans, with between 7 and 72% of the population infected, depending on the geographic area.
Malaria infects an estimated 300 million people, and is spread by mosquitoes, transfusions, and shared hypodermic needles. Control of mosquito populations has led to declines in malaria in many areas. Infected individuals can be treayted with a variety of medicines. However, some of the sporozoans that cause malaria heve developed immunity to some of the more commonly employed medicines.
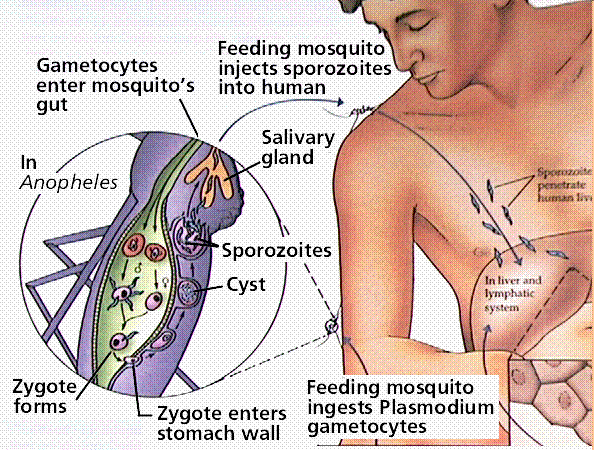
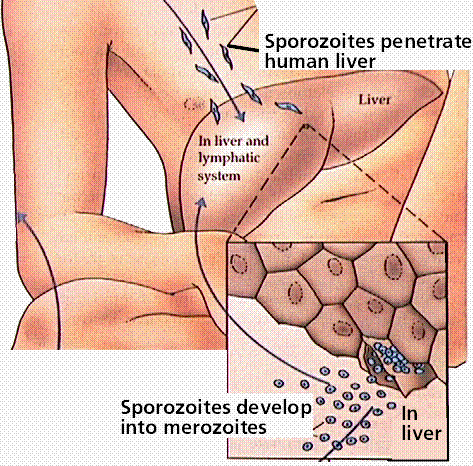
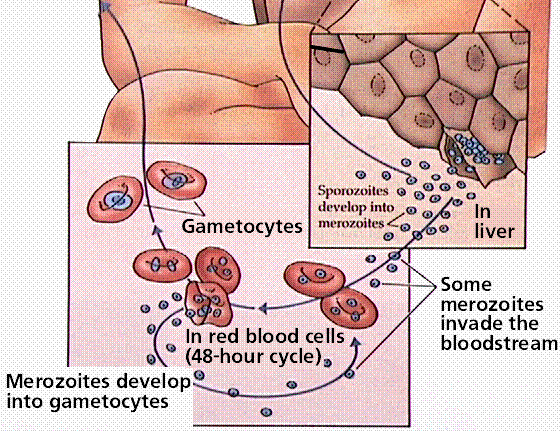
Image from W.H. Freeman and Sinauer Associates, used by permission.
Ciliates are complex, heterotrophic protozoans that lack cell walls and use multiple small cilia for locomotion. To increase strength of the cell boundary, ciliates have a pellicle, a sort of tougher membrane that still allows them to change shape. Most of the 8000 species are freshwater. Most ciliates have two nuclei: a macronucleus that contains hundreds of copies of the genome and controls metabolisms, and a single small micronucleus that contains a single copy of the genome and functions in sexual reproduction. Paramecium is a common ciliate seen by students in introductory biology classes. .
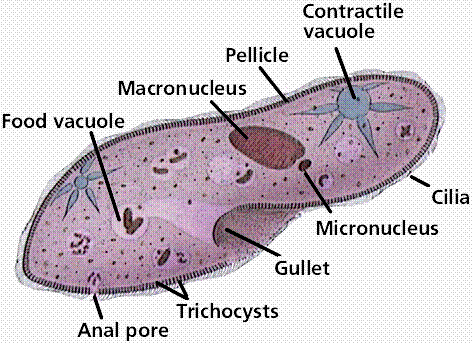
Image from W.H. Freeman and Sinauer Associates, used by permission.
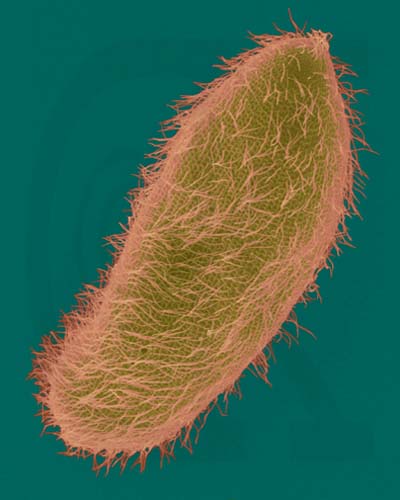
Paramecium multimicronucleatum - Ciliated Protozoan (SEM x1,600). This image is copyright Dennis Kunkel http://www.pbrc.hawaii.edu/~kunkel/gallery, used with permission.
Since ciliates (an many freshwater protozoans) are hypotonic, removal of water crossing the cell membrane by osmosis is a significant problem. One commonly employed mechanism is a contractile vacuole. Water is collected into the central ring of the vacuole and actively transported from the cell
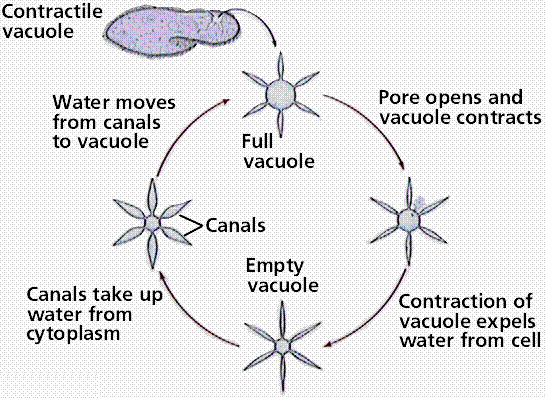
Image from W.H. Freeman and Sinauer Associates, used by permission.
Food is taken into the cell by an oral groove (or gullet), where small particles of the food are phagocytosed into food vacuoles. These travel through the cytoplasm and are digested, with the molecules eventually passing into the cytoplasm.

Image from W.H. Freeman and Sinauer Associates, used by permission.
Ciliates travel along a spiral path, with the cell rotating along its long axis and the direction of travel resembling a sine wave.
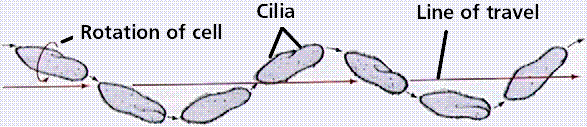
Image from W.H. Freeman and Sinauer Associates, used by permission.
Algae are a polyphyletic group that includes several smaller monophytletic groups. The fossil record of algae dates to the precambrian (possibly the Bitter Springs Formation, 1.2-1.4 billion years old), with undeniable algae appearing during the Paleozoic Era, by about 500 million years ago.
Most algae use photosynthesis at least part of the time. Algae are subdivided by their type of wall, photosynthetic pigments, and method of food storage. Photosynthetic pigments and storage of sugars are quite diverse within the algae. Algae are major components of the phytoplankton, an important source of oxygen and the base of many food webs in the oceans and freshwater. Body styles range from single-celled to colonial (possibly simple multicellular).
Organisms in the Euglenophyta have two flagella, a contractile vacuole, a photoreceptive eyespot, several chloroplasts, lack a cell wall, and can live as autotrophs or heterotrophs. Some autotrophic species of Euglena become heterotrophic when light levels are low.
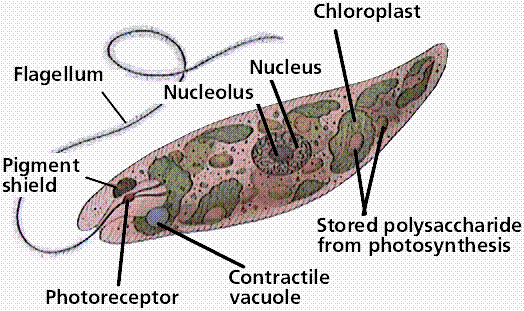
Image from W.H. Freeman and Sinauer Associates, used by permission.
This group of freshwater, marine, and terrestrial algae includes the golden brown algae and diatoms. Food is stored as oils, and photosynthetic pigments include chlorophyll a and c and yellow carotenoid pigments.
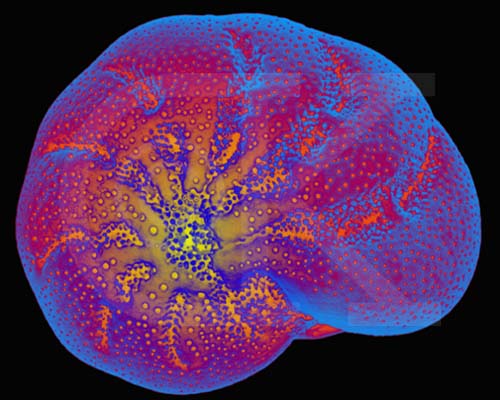
Diatoms secrete a silicon dioxide shell (called a frustule) that forms the fossil deposits known as diatomaceous earth, which is used in filters and as abrasives in polishing compounds. Diatoms divide into two groups,the pennaleans with bilateral symmetry and elongated shape, and another, the centraleans, with radial symmetry and a rotund shape. Certain diatoms also are important indicators of water quality, while others are useful fossils for age-dating Quaternary deposits.


The above images of diatoms (Protista) are from http://WWW.bgsu.edu/departments/biology/algae/index.html.
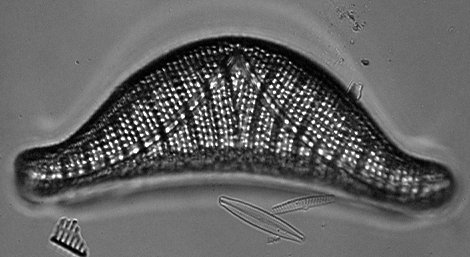
The above differential interference contrast (DIC) image of Epithemia smithii is modified from http://www.umich.edu/~phytolab/GreatLakesDiatomHomePage/Epithemia/Epithemiasmithii/Epithemiasmithii.html.

Centric Diatom Silica Skeleton (SEM x7,220). This image is copyright Dennis Kunkel http://www.pbrc.hawaii.edu/~kunkel/gallery, used with permission.

Saltwater Pennate Diatom Frustule (SEM x4,800). This image is copyright Dennis Kunkel http://www.pbrc.hawaii.edu/~kunkel/gallery, used with permission.
Dinoflagellates have two flagella emerging near the middle of the cell, one girdling the cell's girth, the other trailing parallel to the cell's long axis. Members of this group have cell walls and store excess sugars as starch. Some dinoflagellates also are "armored", having numerous plates that cover the cell. Ornamentation on these plates can be quite beautiful. Certain dinoflagellates live symbiotically inside corals, and are known as zooxanthellae. Most dinoflagellates are autotropic, with chlorophyll a, chlorophyll c, a unique pigment peridinin, and other carotenoids. Some dinoflagellates are heterotrophic. Cell division in dinoflagellates differs from most protistans, with chromosomes attaching to the nuclear envelope and being pulled apart as the nuclear envelope stretches. During cell division in most other eukaryotes, the nuclear envelope dissolves.
The fossil record of dinoflagellates is excellent, with most palynologists accepting fossils from the Triassic as representing some stage of the dinoflagellate life history. Dinoflagellate-like fossils have been reported from the Ordovician, but none in the 200 million year gap between that period and the Triassic. Acritarchs, an abundant group of precambrian and Paleozoic microfossils, may in part be dinoflagellates or might also represent some other group of algae.
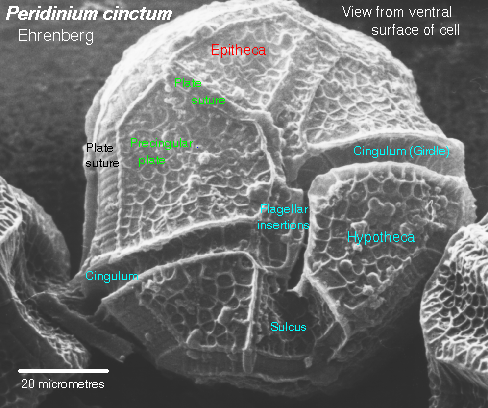
This image of Peridinium is from http://megasun.bch.umontreal.ca/protists/peri/peri.sem2.lab.gif.
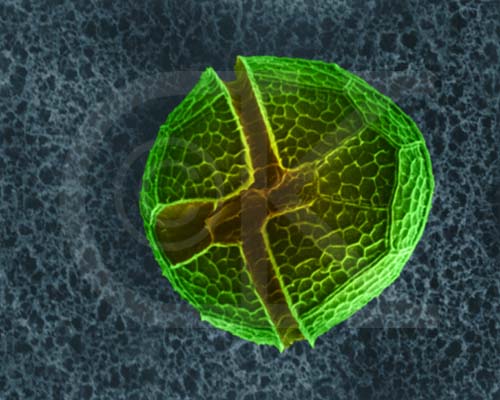
Dinoflagellate, Peridinium sp. (SEM x3,500). This image is copyright Dennis Kunkel http://www.pbrc.hawaii.edu/~kunkel/gallery, used with permission.
Red tides are caused by population explosions of certain dinoflagellates that release a neurotoxin into the environment. Shellfish concentrate this toxin and it can kill people who eat the contaminated shellfish. Woods Hole Oceanographic Institute maintains a nice page describing What are Harmful Algal Blooms (HABs)?

This image of a red tide is from http://www.redtide.whoi.edu/hab/rtphotos/noctiluca.jpg.
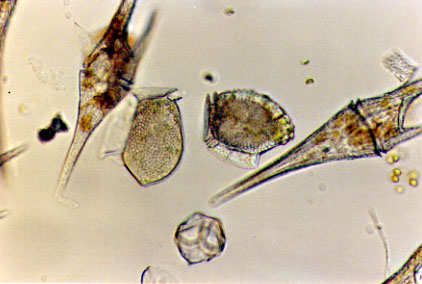
The dinoflagellates from this bloom are from http://www.redtide.whoi.edu/hab/rtphotos/dinophysis.jpg.
Fish kills have been linked to the dinoflagellate Pfiesteria.
Red algae have large amounts of the red pigment phycoerythrin, range from unicellular to multicellular, sometimes more than one meter in length. Red algae are thought to have originated by symbiosis of cyanobacteria (which also have phycoerythrin). Some red algae are important contributors to tropical reefs. Carrageenan is an additive to puddings and ice creams; dried sheets of red algae are used in some Japanese dishes.


The above left image of a piece of the red alga Erythrophyllum delesseriodes is from http://www.sonoma.edu/biology/algae/Erythrophyllum.html. The image on the right is Microcladia coulteri from http://www.sonoma.edu/biology/algae/Microcladia.html.
Brown algae are multicellular, have the accessory pigment fucoxanthin (a brown pigment), and include the giant kelp that can be over 100 meters long. Brown algae are used in foods, animal feeds, and fertilizers and as a source for alginate, a chemical emulsifier added to ice cream, salad dressing, and candy.
Fucus is a brown alga differentiated into a floating "blade", flotation bladder, stalk (or stipe) and basal holdfast. Sargassum, common in the Sargasso Sea region of the Atlantic Ocean, floats and maintains position by a flotation bladder filled with gas.

The above image of Fucus is from http://www.ucmp.berkeley.edu/chromista/phaeophyta.html.

The above image of Nereocystis is from http://www.sonoma.edu/biology/algae/Brown.html.
Green algae have cellulose cell walls, chlorophyll a and b, store excess photosynthetic product as starch, and have long been considered the undoubted ancestors of plants. Body types in the green algae include unicellular to colonial and simple multicelluar.
Chlamydomonas and similar cells appear to be a starting point within this group. Autotrophic, unicellular forms with a single, cup-shaped chloroplast and two apically inserted flagella, these small cells also possess a contractile vacuole and pyrenoid. Excess sugars are stored as starch surrounding the pyrenoid.
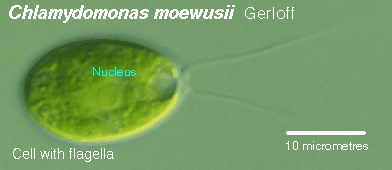
This image is from http://megasun.bch.umontreal.ca/protists/chlamy/chlamy21.lab.gif.
Multicellular green algae have some division of labor, producing various reproductive cells and structures. Ulva, the sea lettuce, exhibits alternation of generations, producing free-living gametophyte and sporophyte forms. The common sea lettuce is usually haploid (the gametophyte) and reproduces asexually. Gametes are produced by mitosis, fuse, and produce a diploid zygote. The 2n zygote germinates and grows to become the sporophyte. Meiosis occurs in certain of the cells in the sporophyte, producing haploid swimming spores that will settle to the ocean floor and produce the next generation haploid gametophyte stage.
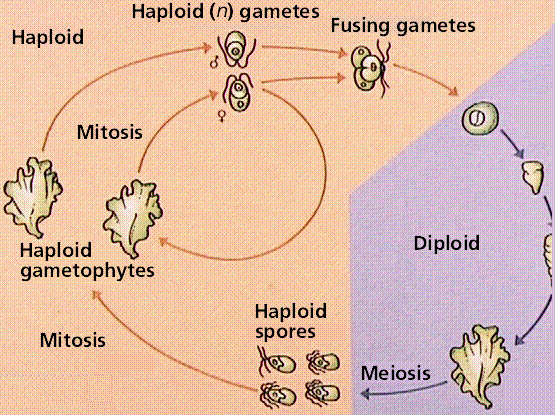
Image from W.H. Freeman and Sinauer Associates, used by permission.
Filametous algae produce gametes by mitosis within one cell of the filament. These gametes are released, fuse to form a diploid zygote that soon undergoes meiosis to produce hapoid zoospores that swim, rest on the sea floor and develop into the next generation gametophyte phase.
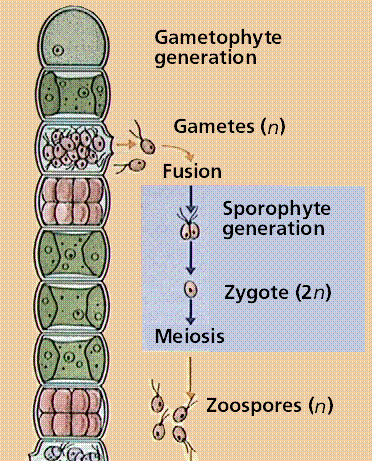
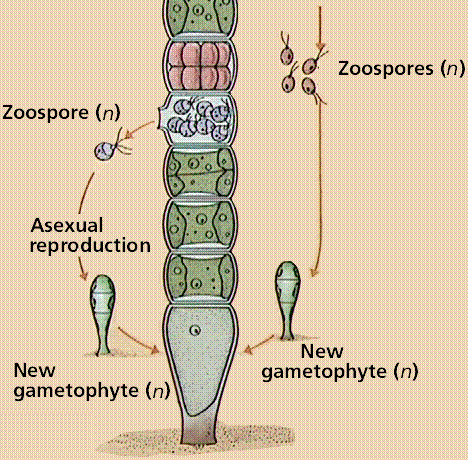
Image from W.H. Freeman and Sinauer Associates, used by permission.
Slime molds are often classified as fungi, although now most specialists consider them a group of protistans. They are not closely related to other groups of plants or animals. Slime molds, which spend part of their life as single-celled forms, can aggregate to form multicellular forms. They thus may represent a transition between unicellular and multicelluar forms, the second major advancement after the evolution of eukaryotic cells.


Image (L) of Physarum, a plasmodial slime mold is from http://www.ucmp.berkeley.edu/protista/slimemolds.html. Image oif sporangia of Physarum from gopher://wiscinfo.wisc.edu:2070/I9/.image/.bot/.332/Slime_molds_M_Ac_sl_so/SLime-PH_Physarumsporangia.
All text contents ©1995, 1999, 2000, M.J. Farabee. Use
for educational purposes is encouraged.
Email: mj.farabee@emcmail.maricopa.edu![]()
Last modified:
2000/01/09:10:01:00
The URL of this page is:
gened.emc.maricopa.edu/bio/BIO181/BIOBK/BioBookDiversity_3.html
{kind=link}
{kind=link}
{kind=link}
{kind=link}