Botany online 1996-2004. No further update, only historical document of botanical science!
Geotropism or Gravitropism

From the beginning of germination display roots a tendency to grow downwards, while shoots grow
upwards (anisotropic growth). A germinating seed can be turned
upside down several times and the root will still start to bend downwards. This behaviour is
another example for a tropism, a movement triggered by a
stimulus. The question is: what is the controlling stimulus?
"It is not the soil’s humidity that causes the direction of the root,
since the root will grow downwards and the shoot will grow upwards even if the plant is placed
in an earth-filled tube where the upper part is moist and the lower is dry. If the plant is put
into a water-filled tube the lower part of which is exposed to light while the upper is darkened,
the direction of growth will still not change: a proof that it is independent of light."
(A. de CANDOLLE, 1834/38)
In 1806 showed the British physiologist A. KNIGHT in a decisive experiment
that the direction of root growth is controlled by gravity. He fixed seedlings on a vertically
standing wheel that was turning around its axis due to a small millwheel fuelled by a fast-flowing
stream. The rotation caused a centrifugal force. The effect caused by this force was also observed
when the wheel was in a horizontal position: at a fast rotation grew all roots outwards
(in the direction of the centrifugal force), at a low rotation did they grow in an angle
of about 45° (the result of both centrifugal force and gravity).
KNIGHT thought at first that the root tips where pulled downwards by
their own weight, but this assumption was soon refuted, since the downward movement occurs
also when the weight of the root tips was compensated for by an opposite weight (JOHNSON, 1828).
A. B. FRANCK introduced the term geotropism in 1868, set it of against
phototropism, and distinguished three types:
 positive geotropism (like in the example just given)
positive geotropism (like in the example just given)
 negative geotropism, and
negative geotropism, and
 transversal geotropism.
transversal geotropism.
Positive and negative geotropism
together are also
called ortho-geotropism. It causes the vertical (orthotropic) orientation of the plant’s axis.
Shoot growth is mostly negatively geotropic since shoots grow upwards even in complete darkness.
Phototropism can therefore be understood as a secondary process, usually of the same direction
as the negative geotropism. Transversal geotropism
is a direction of growth that is vertical to the shoot axis. The direction of many lateral
shoots, side roots, leaves, etc. is described by this term. All directions diverting from
that of the shoot axis are called plagiotropic. Orthotropic organs (shoot axis, main root)
are usually of radial symmetry, plagiotropic organs (leaves, side roots, etc.) are mostly
of dorsiventral organisation.
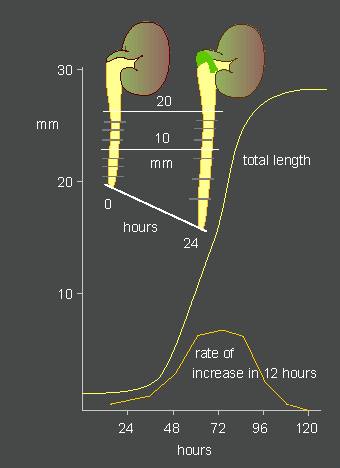 J. v. SACHS extended the experiments of KNIGHT and supplemented them by the invention of
the clinostat (1879) that helped him to stop the bending (compensation of gravity by the
centrifugal force). By marking the single parts of the root with water colour marks (today
exchanged against other types of labelling) did he show that in the root,
too, it is not the
tip itself that elongates but the section above (a growth zone). Plant anatomical studies
proof that the elongation of one side is caused by an elongation of the respective cells.
J. v. SACHS extended the experiments of KNIGHT and supplemented them by the invention of
the clinostat (1879) that helped him to stop the bending (compensation of gravity by the
centrifugal force). By marking the single parts of the root with water colour marks (today
exchanged against other types of labelling) did he show that in the root,
too, it is not the
tip itself that elongates but the section above (a growth zone). Plant anatomical studies
proof that the elongation of one side is caused by an elongation of the respective cells.
We have not yet answered the questions how the stimulus is perceived and how it is transmitted.
In 1892 postulated F. NOLL (1858 – 1908, professor at Bonn, later at Heidelberg) the existence
of microscopically small, mobile components in the cells of the root tip exerting a pressure
on the plasma of the bottom side of the cell. The stimulus produced by the local pressure would
then have to be converted into a stretching of the cell and into an increased growth by the
cell’s plasma. The plant anatomists B. NEMEC at Prague and the Austrian G. HABERLANDT who
regarded the central cells of the calyptra as the location for the mobile particles
following gravity were able to confirm this suggestion
(statolith theory). They could identify the particles
as amyloplasts.
NOLL’s suggestion remained insecure in one point, since the relocation
of the statoliths does not cause a stretching of the starch-containing cells’ walls, but
an elongation of the cells in the growth zone. The question for stimulus forwarding is inevitable.
A definite answer has not been given yet, though it is known that auxine participates.
Its distribution pattern is not identical with that observed in phototropism, though.
The following observations indicate its participation:
The amyloplasts’ mobility decreases from tip to base just like the
influence of gravity upon the auxin transport. Strong centrifugal forces cause relocations
of the amyloplasts and simultaneous relocations of auxin in the basal parts, too.
Mutants with smaller amyloplasts display a smaller rate of relocation,
less influence of gravity upon the auxin transport, and less geotropic bending. The latter
does not mean that the mutant has a generally reduced ability to bend since the phototropic
reactions of mutant and wild type do not differ.
When talking about phototropism did we mention the behaviour of the Phycomyces sporangium.
A comparable example is known for geotropism, too. It is the reaction of the single-celled rhizoids
of the stonewort Chara. They are transparent, indifferent towards light, and the movements
of their particles can be easily observed with a microscope. The existence of strongly refractive
cellular inclusions ("Glanzkörper") in the tip of the rhizoid the
relocation of which correlates with a geotropic bending is especially striking (J. BUDER,1961).
Based on these observations tried A. SIEVERS and his collaborators (Botanical Institute of the
Universität Bonn) to make out a direct correlation between the relocation of the
"Glanzkörper" and the geotropic bending by microscopical,
electron microscopical, and biochemical analyses. Experiments with cytochalasin B
that stops the activity of microfilaments showed that the relocation of the statoliths
is an actively controlled process involving microfilaments. Electron microscopical images
showed that Golgi-vesicles participate in the cellular growth, and that they are moved to
those areas where cellular elongation takes place as soon as bending starts. In summary,
one can draw the following conclusions:
|
Statoliths hinder cellular growth in the areas where they are
close to the plasma (exactly the opposite to what NOLL postulated). In places where the
statoliths sediment at first do they cause slight denting in.
|
 |
Golgi-vesicles gather at the side opposite to the statoliths
where they participate in the beginning synthesis of the cell wall that leads finally
to bending. By putting weight on the root tips can the growth rate of the single sections be
exactly monitored.
|
 |
If statoliths are relocated to a vertically growing rhizoid,
is the regular longitudinal growth inhibited.
|
Even though the observations made with Chara are fairly impressive,
do they not explain the causal chain in multicellular roots where stimulus perception and
gravitropic reaction take place in different cells even separated by other, non-reactive cells.
The findings do stress another point, though: they show that the black box between stimulus
perception and reaction can be assembled from different system components.
© Peter v. Sengbusch - Impressum