Botany online 1996-2004. No further update, only historical document of botanical science!
Protoplasts
 |
At the beginning of the sixties used E. C. COCKING (University
of Nottingham) an enzyme preparation for the degradation of cell
walls and yielded thus single protoplasts from tomato root tips,
though it took until 1968 before I. TAKEBE et al. (Institute
for Plant Virus Research, Aobacho, Chiba/Japan) developed a technique
for the production of large amounts of active protoplasts from
mesophyll cells of Nicotiana tabacum that became standard.
It spread fast due to its simple use, and within short time succeeded
a number of laboratories in the yield of protoplasts from tissues
of different plant species. Protoplast
preparation was at first done in two steps:
|
- At first was the middle lamella dissolved by pectinases,
- then was the cell wall broken down by cellulase.
The enzymes required for this procedure are usually not pure,
but crude extracts from certain bacteria and fungi. Nowadays is
the method often shortened to just one operation in which a mixture
of enzymes is applied. Due to the high osmotic pressure within
the protoplast are isotonic media used. They contain usually mannitol
and/ or sorbitol. Besides contains protoplast media a variety
of inorganic salts and several vitamins. It facilitates the maintenance
of protoplasts capable of dividing for several days. They can
be cultivated on agar dishes where they grow into small visible
calli.
Among the most striking properties of active protoplasts is the
fast regeneration of a cell wall that does always precede protoplast
division. It seems as if the development of a cell wall was a
causal precondition of both cell division and development of polarity.
Polarity and positional information are lost during protoplast
generation. Both have to be developed anew, if a new plant is
going to regenerate.
Protoplasts can be used for the production of 'vacuoplasts' (pure
vacuoles) and subprotoplasts that consist of nucleus and plasma
alone. The single fractions can be separated from each other and
concentrated by density gradient centrifugation. What is the use
of protoplasts?
- Protoplasts can regenerate into complete plants.
- Protoplasts of the same or different origin can fuse with each
other. The fusion product may even regenerate into a plant. This
is also called somatic hybridization.
- The protoplast can take up macromolecules (nucleic acids and
proteins), viruses, cell components like chromosomes and chloroplasts
by phagocytosis.
- Protoplasts are suitable for the study of the molecular architecture
of plant cells
Regeneration
I. TAKEBE, G. LABIB and G. MELCHERS (Max-Planck-Institut für
Biologie, Tübingen) regenerated complete plants from tobacco
protoplasts in 1970.
Y. Y. GLEBA (Academy of Science of the Ukrainian SSR, Kiew) developed
a technique for the cultivation of isolated protoplasts in micro-droplets.
A number of thus cloned protoplasts regenerated into complete
plants.
By now have protoplasts from a number of dicotyledonous angiosperms
been regenerated into fertile plants. Solanaceae pose the smallest
problems. In contrast regenerated protoplasts from leguminosae
or monocots into complete plants only after the development of
costly procedures that do still not always succeed. Regeneration
requires an addition of phytohormones to the medium. It looks
as if the mesophyll cells of monocots would loose the respective
receptors. This may also be the reason why hormone-like herbicides
impair dicots but not monocots (cereals). On the other hand is
the lacking ability to regenerate into protoplasts of economically
important cultivated plants a large handicap when trying to transfer
the knowledge gained with this method to the agricultural practice.
Fusion of Protoplasts - Which Part Play Species Boundaries?
The aggregation of two or more protoplasts is not enough to start
a fusion. Protoplast surfaces bear strong negative charges. In
contrast to animal cells is the surface charge not caused by sialic
acid residues but by phosphate groups. Intact protoplasts in suspension
do thus repel each other. They can be very impressively linked
and fused by the addition of calcium ions or polyethylenglycol.
Electric fields are an alternative.
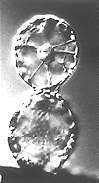
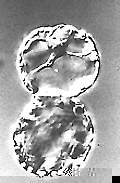

Fusion of Protoplasts in an Electric Field (electrofusion;
H.-U. KOOP, H.-G. SCHWEIGER, 1985)
Sexual hybridization occurs when haploid cells generated in a
previous meiosis fuse. The fusion of somatic diploid cells should
generate a tetraploid fusion product provided that the nuclei
fuse, too. If this is the case, it is spoken of a synkaryon.
A fusion product where the nuclei stay separate is called a heterokaryon.
For conditions comparable to sexual hybridization is haploid starting
material needed. S. GUHA and S. C. MAHESHWARI (Botanical Institute
of the University of Delhi, 1966) developed a medium for the cultivation
of anthers from Datura innoxia. The anthers grew into seedling-like
plantlets also called embryoids.
These descendants of meiosis that did not develop further into
pollen turned out to be 'plants out of gones'. They had not the
diploid number of chromosomes normal for plant tissues but only
a haploid set. Gones is the name for the (haploid) products of
meiosis (independent of the sex). By now have haploid plants from
a whole number of different plant species, both mono- and dicots,
been cultivated successfully. They are used as starting material
for the production of protoplast.
In order for haploid plants to propagate by seeds do they first
have to become diploid. This is achieved rather easily since colchicine
was found out to be very suitable for this task in1937. Diploid
plants thus generated have the advantage of being completely homozygous.
Haploid species of Nicotiana (N. tabacum, N. sylvestris)
have first been used for intraspecific, later on also for interspecific
protoplast fusion. The plants yielded by regeneration of such
fusion products do not differ from sexually generated ones. Fusion
products of parental varieties that complement their chlorophyll
defects are especially suitable. They can be recognized by their
green colour (G. MELCHERS and G. LABIB, Tübingen 1974).
Interspecific fusions can be
generated rather easily. It is even possible to fuse plant protoplasts
and animal cells (fibroblasts) and to keep the fusion product
alive for several hours. Such experiments are not performed for
regeneration purposes but in order to understand the co-operation
of membranes or the translation of single plant genes within the
animal cell's plasma. With few exceptions regenerate interspecific
fusion products only, if sexual hybridization is successful, too.
Interspecific heterokaryons, especially those of not closely related
species, do not form synkaryons. Nuclear division is asynchronous
and as a consequence are chromosomes lost: the whole system looses
its balance. Despite this have some somatic interspecific hybridizations
been successful. G. MELCHERS succeeded in 1978 in the fusion of
tomato and potato protoplasts. The fusion products (two German terms: meaning Tomate x Kartoffel or Kartoffel x Tomate = "Tomoffel" or "Karmate") regenerated
into whole plants that flowered but did not develop fertile seeds.
The fusion product of Datura innoxia and Datura stramonium
developed by O. SCHIEDER (Max-Planck-Institut für Züchtungsforschung,
Köln) did even produce fertile seeds. It can therefore be
regarded as a new species: Datura straubii. Datura
- species are used for the production of medically important alkaloids
hyoscyamine and scopalamine. Datura
straubii has a larger growth capacity and a 20 - 25 percent
higher alkaloid content than its two parental species.
Absorption of Macromolecules, Viruses, and Organelles
Protoplasts can absorb foreign molecules and organelles by phagocytosis.
This furnished proof that one cell can be infected by more than
just one virus particle and that different strains of viruses
can multiply within one cell. Furthermore were protoplasts transformed
with Ti-DNA from Agrobacterium tumefaciens which made the
system interesting for genetic engineering. The absorption of
plastids permits an analysis of the co-operation between the nuclear
and the plastidic genomes. Absorbed foreign genes can, too, make
up for defects within the protoplast genome. The fusion with inactivated
protoplasts of Physalis and Datura transferred a
nitrate reductase activity to a mutant of Nicotiana tabacum
with a deficient nitrate reductase. Transformation proved to be
stable (R. P. GUPTA, M. GUPTA, O. SCHIEDER, 1982).
The use of modern analytic techniques like gel electrophoresis,
evidence of enzyme activities within a gel or isoenzymes verifies
the success of fusion experiments. Ribulose - 1,5 - bisphosphate
carboxylase has an important marker function since it consists
of nuclear and plastidic encoded subunits. At the same time offers
the analysis of this protein information about the activities
of the plastids and nucleus of the parental species within the
fusion product.
After protoplast techniques had been established became the production
and subsequent isolation of mutants important. Auxotrophic strains,
i.e. strains that are dependent on the supply of certain nutrients
were of special interest. Only few successes were achieved, one
reason of which may be that a large part of angiosperm genomes
are allopolyploid and that genetic information exists in several
copies even in haploids. This may easily compensate for a defect
gene. In addition are plants able to produce one and the same
product by different metabolic pathways. A possible way out of
the dilemma could be the use of 'monohaploid' stock plants (1
x instead of 1 n). In the case of cultivated plants would that
mean that their original varieties have to be found first.
The Study of the Molecular Architecture of Plant Cells
S. M. WICK et al. (Australian National University, Canberra,
1981) produced protoplasts from pre-fixed cells since the original
cell shape is lost during protoplast generation from native cells.
The thus generated protoplasts are dead, but they have kept their
shape, and indirect immunofluorescence allows the depiction of
the cell's structural elements' distribution. The use of lectins
showed that the protoplast surfaces of different species carry
different carbohydrate patterns and that the lectin-binding properties
of the plasmalemma differ fundamentally from those of the intracellular
membranes and the tonoplast.
© Peter v. Sengbusch - Impressum