Botany online 1996-2004. No further update, only historical document of botanical science!
The wave length problem - stabilization of a monotone gradient by a feedback on the source density
How to generate structures close to each other, how at a distance: head, foot and tentacle formation in Hydra
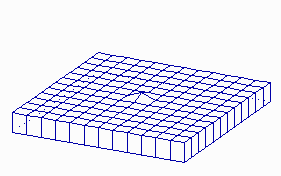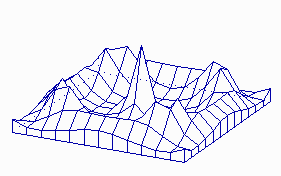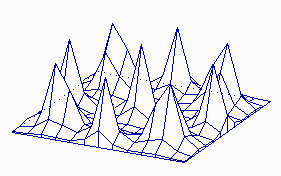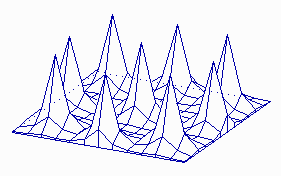
Fig. 4: A more or less regular arrangement of activator peaks results in a field that is large compared to the range of the inhibitor.


Fig. 5: Regular periodic structures emerge if the pattern formation works during growth. (a) an alternating (distichous) pattern of leaf initiation is simulated by assuming a cylinder that grows at its upper end. New maxima appear in 180° arrangement. (b) If the diameter of the cylinder is somewhat larger and/or the range of the inhibitor is shorter, pairs of maxima at opposite sides. Each subsequent pair is displaced by 90° (decussate pattern). The existence of helical arrangements of leaves suggest a more complex mechanism (see Fig. 19-23 ).

Fig. 6: Stripe-like distributions result if the autocatalysis saturates.
Patterns that are periodic in space are formed if the size of the field is larger than the range of the inhibitor. In a sheet of cells maxima with more or less regular spacing can be formed (Fig. 4). This is appropriate for the initiation of periodic structures like bristles or stomata. The pattern becomes more regular if the pattern forming reaction works already during growth. In Fig. 5 pattern formation is shown on a cylinder that grows by proliferation at it upper end. New maxima appear during growth at regular distances on alternating sides. This is a simple model for the initiation of leaves on a growing shout. Stripes, a pattern frequently encountered in development, can be formed if the rate of activator autocatalysis saturates at high activator concentrations (Fig. 6). This leads also to a limitation of the inhibitor production. More cells become activated at a lower level until sufficient inhibitor is produced. In other word, the activated regions have the tendency to enlarge, However, in order to become activated, a close neighbourhood to non-activated cells is essential into which the inhibitor can be dumped. Both requirements, large activated patches and a direct neighbourhood of non-activated cells, seems to contradict each other. This is, however, not the case. In a stripe-like activation pattern, each activated cell has an activated neighbour and non-activated cells are close by. Stripe formation is a very frequent phenomenon at very different developmental situations. Transitions between patch- and stripe-like pattern can be frequently seen of the skin pattern of tropical fishes. Kondo and Asai (1995) have observed the dynamic regulation of stripes of pigmentation on growing tropical fishes.
Usually the size of a morphogenetic field increases during the growth of the embryo. In a usual reaction-diffusion system a graded concentration profile can be maintained only over a range of about a factor two. With increasing field size, a tendency exists to change from a monotone distribution into a symmetric and ultimately into a periodic distribution either by insertion of new (Fig. 5 or by splitting of existing maxima. This is inappropriate if the graded concentration should be used in the growing embryo as positional information for the determination of the primary body axes. Multiple maxima would lead, for instance, to several heads instead of one.
Some basic biological observations provide important hints of how this wave length problem has been solved by nature. Hydra maintains its polar structure over substantial growth but, nevertheless, a fragment 1/10 of the normal body size is still able to regenerate. In such a fragment the regeneration of the head occurs always at the side pointing towards the original head. Morgan (1904) interpreted similar observations with Tubularia by assuming that a systematic change in the ability for head regeneration exists and that a competition takes place. The tissue originally closer to the head has a head start and wins the competition. The relative position is decisive.
In terms of our model, the range of dominance of the activated over the non-activated region (the so-called apical dominance) can be increased by an order of magnitude if a activator or the inhibitor has a feedback on the ability of the cells to produce the pattern forming reaction (green in Figs. 7, 8; Meinhardt and Gierer, 1974). Due to this feedback, the source density becomes graded too. In a region of low source density, the initiation of secondary maxima is unlikely. In contrast, the rising source density at an existing maximum stabilizes this maximum. Thus, the polar distribution becomes stabilized. The graded source density provides the long-lasting information about the polarity of the system. A small fragment regenerates a pattern according to the original polarity since the graded source density provides a systematic headstart for some cells to outcompete the others. Since the source density has a long time constant, it remains essentially unchanged during pattern regeneration (see Fig. 8). Although secondary maxima are successfully suppressed, the regeneration is not impaired. Regeneration can be fast since no symmetry breaking and no communication by diffusion over the total field is required.
Since the source density concept is very important to explain many biological observations, the mechanism should be explained with a somewhat anthropomorphic analogy. A king (or any other local hero) has usually a strong tendency to suppress other's from becoming a king too - a long range inhibition. On the other hand, he promotes individuals in his surrounding to obtain different levels in a hierarchy, to become ministers etc. In this way, the centre of power generates a hierarchy. If the centre of power would become void, due to this non-uniformity is usually clear who will win the subsequent competition.

Fig. 7: Pattern formation in Hydra: generation of complex patterns by linkage of several pattern forming reaction. Primary head (blue) and foot activation (black) appear at opposite end of the field due a coupling via the source density (green). Tentacle activation (red) appears close to the hypostome since it requires a high source density but it is locally suppressed by head activation.
The complexity of the patterns in higher organisms requires a hierarchical linkage of many pattern forming reactions. One or more patterns generate the precondition for a subsequent pattern. The combinatorial possibilities are very large, making modelling very difficult. That nevertheless the modelling of complex patterning processes is possible should be illustrated with a model for the freshwater polyp Hydra (Fig. 7, Meinhardt, 1993).
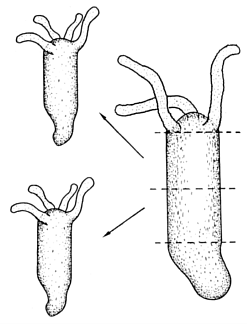
Hydra is under control of two organizing regions located at opposite ends, the head and the foot. This is common of many morphogenetic fields. How can it be achieved that two organizing centres reliably appear at opposite positions of an extended field? For Hydra a simple cross-inhibition is not appropriate since in small (young) animals head and foot must appear very close together. If at such short distances a mutual inhibition between the head and the foot system would be at work, this would lead to a suppression of the foot by the nearby head or vice versa. This problems disappears when the spacing between the head and foot system is achieved by an interaction via the source density. As mentioned above, the head activation appears at the position of the highest source density. If the foot system has the opposite behaviour, i. e., it appears at the lowest source density, the foot is formed at the maximum distance from the head (Fig. 7). Nevertheless, head and foot system can coexist at a close neighbourhood in small animals since no direct inhibition is involved. The graded source density only generates a preference. Experimental evidence indicates that the foot also lowers the source density, contributing in this way to the maintenance of the source density gradient.
Many structures emerge during development close to each other in a precise arrangement. A controlled neighbourhood of structures is enforced if one structure activates the other on long range but excludes it locally (Meinhardt and Gierer, 1980). In Hydra, the tentacles appear around the hypostome, the opening of the gastric column. Many experiments can be accounted for by the assumption that tentacles are under control of a separate activator-inhibitor system that also depends on the source density. Since the source density increases under the influence of the primary head system, the latter generates the precondition for tentacle initiation. Locally, however, the head signal suppresses tentacle formation. Thus, tentacle formation (red in Figs. 7, 8) occurs next to the head.
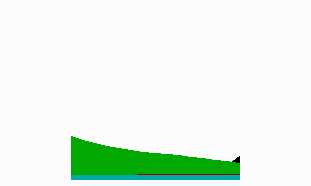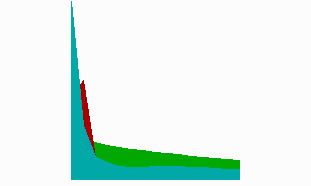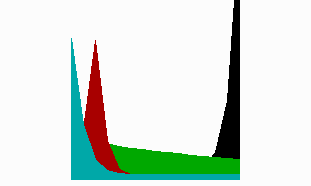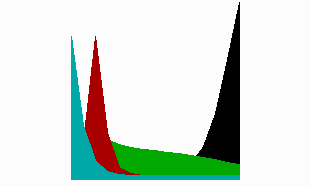
Fig. 8: In regenerating near-head fragments, the generation of the tentacle signal (red) precedes head activation (blue). It occurs first at the tip and shifts later to its final position, in agreement with the experimental observations (after Meinhardt, 1993).
The model accounts for a strange-appearing observation. With tentacle-specific antibodies, Bode et al. (1988) have shown that after head removal, tentacle activation first reappears at the very tip of the gastric column. It is only later that this activation becomes shifted to the position where the tentacles eventually appear. Since the tentacles are formed close together, the tentacle inhibitor need not to diffuse very far. In terms of the model, the tentacle inhibitor can have a short half life. Thus, after removal of the head and the tentacles, the tentacle inhibitor fades away more rapidly than the head inhibitor. Therefore, the tentacle activator can reappear sooner than the head activator. Since no suppressing head activator is present, this happens at the highest possible source density (green in Fig. 8), i.e., at the front end of the remaining gastric column. After the trigger of the primary head activation at the same position, tentacle activation becomes shifted to the final location. This shift after the trigger of the head signal (blue) is clearly visible in the simulation. The prediction was further that the sequence of events is the reverse after cutting closer to the budding region, a prediction that has been meanwhile verified (Technau and Holstein,1995). In this situation, first the formation of the head signal takes place. The formation of the tentacle signal occurs afterwards at the correct position.