Botany online 1996-2004. No further update, only historical document of botanical science!
The characteristic biosynthetic pathway of green plants is that of carbohydrates via photosynthesis. Because of their importance and the complexity of the involved processes, we will deal with it in an extra section.
For our present purpose, it is enough to know that glucose and other phosphorylated sugars are generated in this process. As we have seen, glucose is by no means just degraded but it serves also as a starting compound of, for example, the biosynthetic pathways of many polysaccharides that are on one hand structural elements of the plant cell (cellulose, for example, as a plant cell wall component) and on the other hand storage compounds (starch). Moreover, plant cells contain a broad range of different but structurally very similar sugars. In this context, the questions how they are converted into each other and how the selectivity of the single reactions is safeguarded are of no small interest.
If not already activated, sugars have, like all other starting compounds of biosynthetic pathways at first to be activated. This occurs either by phosphorylation or by binding of a sugar residue to a nucleotide.
Fructose-6-phosphate is an important intermediate of both photosynthesis and glycolysis. It is at equilibrium with glucose-6-phosphate and this again is at equilibrium with glucose-1-phosphate.
Glucose-1-phosphate and UTP react to UDP-glucose that again polymerizes with fructose-6-phosphate to form saccharose phosphate. Upon cleavage of the phosphate, saccharose is produced. The reaction takes part in chloroplasts.
Starch is generated by coupling glucose-1-phosphate to adenosine diphosphate
Glucose-1-phosphate + ATP + H2O > ADP-glucose + 2 Pi.
This means that the linkage to a nucleoside diphosphate is decisive for the taken pathway.
UDP-glucose (but not ADP-glucose) can also be incorporated into glycolipids and glycoproteins.
Two enzymes are necessary for starch production: one for the start and the chain elongation (alpha 1 > 4 glycosidic linkage) with monosaccharid units the other for the introduction of alpha 1 > 6 glycosidic linkages as well as for linking together different chains (via alpha 1 > 4 glycosidic bonds). This enzyme does not couple single glycosyl units to existing chains but transfers whole chains. It is distinguished between intramolecular and intermolecular transfer.
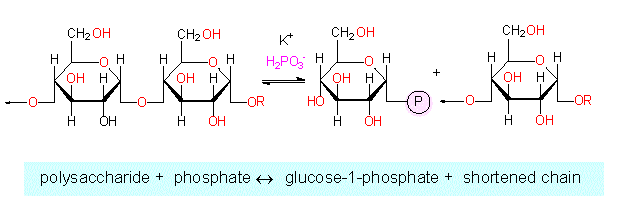
At least four different enzymes are needed for the breakdown of starch. Two ways exist:
The hydrolytic breakdown by an amylase. The starch is broken down to disaccharide units (maltose). A degradation next to points of branching is impossible. Dextrin residues remain.
The phosphorylytic degradation by a phosphorylase. For this pathway, inorganic phosphate is indispensable. The final product is glucose-1-phosphate. In contrast to the amylase reaction, this one is reversible.
Conversion of One Sugar into Another by Epimerization:
The conversion of glucose to galactose is an example. Starting compound is again a glucoysl residue bound to an UDP. NAD+ is also necessary though it remains unaltered in the overall reaction.
UDP-D-glucose + NAD+ > UDP-4-keto-D-glucose + NADH + H+UDP-4-keto-D-glucose + NADH + H+ > UDP-D-galactose + NAD+
Generation of C5-sugars or the
Oxydative Pentose Phosphate Cycle. The oxydative pentose
phosphate cycle is in many respects the conversion of the reductive
pentose phosphate cycle that we know better as the Calvin
cycle. We will delve into it in more detail when dealing with
photosynthesis.
Contrary to the Calvin cycle that takes place in chloroplasts, the reactions of the oxydative pentose phosphate cycle take place in the cytosol. Also, its reactions are not confined to plant cells alone. By oxidation of glucose-6-phospsphate and splitting off of carbon dioxide, ribose-5-phosphate is generated. The surplus electrons are transferred to NADP+.
glucose-6-phosphate + 2 NADP + H2O > ribose-5-phosphate + 2 NADPH + H+ + CO2.
It seems unnecessary to point out the importance of ribose-5-phosphate since it is an essential part of ATP, CoA, NAD+, FAD, RNA, DNA and others.
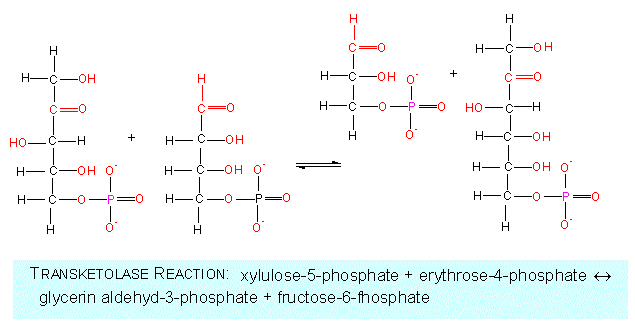
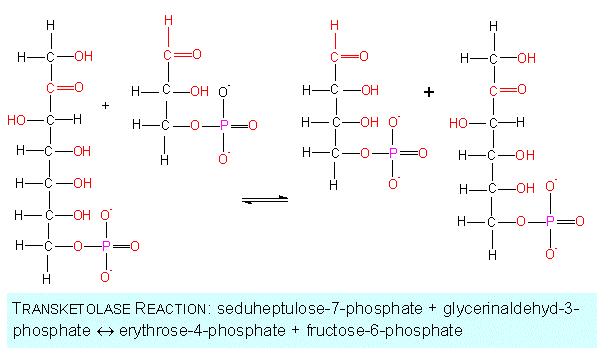
Among the further reactions are the conversions to C3-, C4-, other C5- and C6- as well as to C7-sugars. The participating enzymes are both transketolases and transaldolases.
Several reactions lead to intermediates that are also
intermediates of glycolysis. They represent links between
the pentose phosphate cycle and glycolysis. Two
(phosphorylated) pentoses, for example,
(xylulose-5-phosphate and ribose-5-phosphate) react to
glycerine aldehyde-3-phosphate and a sugar with seven
C-atoms, the seduheptulose-7-phosphate. In the reaction,
a ketogroup is transferred from one sugar to another
intermediate of glycolysis.
On the other hand, the reaction can be reversed by a transaldolase reaction generating erythrose-4-phosphate and a fructose-6-phosphate. The latter is an intermediate of glycolysis, too, while the first is, as we have already seen, necessary for the synthesis of the amino acids serine and glycine.
Which of the single pathways is chosen depends mostly on the supply of ATP, NAD+, NADP+ and others. The supply and the need for them decides whether an intermediate remains in the pentose phosphate cycle or is fed into glycolysis to be degraded. The coupling of both pathways offers also the opportunity to regain glucose. When discussing glycolysis we saw that two of its reactions are irreversible, i.e. glucose cannot be regained by simply changing the direction of the glycolytic reactions. But it can be regained if an intermediate (glycerine aldehyde-3-phosphate) is fed into the oxidative pentose phosphate cycle and the equilibrium is such as to regain glucose.
 ribose-5-phosphate
ribose-5-phosphate