Botany online 1996-2004. No further update, only historical document of botanical science!
There exist two techniques for the depiction of the structural organization of epidermal surfaces. One is scanning electron microscopy, the other the microscopic analysis of imprints in synthetic material. Both techniques are demonstrated by a number of examples.
The epidermis has numerous functions:
|
|
protection against various chemical and physical influences, against being fed upon by animals and against infestation by parasites |
|
|
protection of the plant against desiccation |
|
|
participation in gas exchange, in secretion of metabolic compounds and in absorption of water |
|
|
site of receptors for light and mechanical stimuli that help to transform signals from the surrounding to the plant |
The epidermis has accordingly a number of differentiated cell types to serve the various functions.Variations typical for certain species and different organizations of the epidermis in the miscellaneous plant organs add to the number of different cells. Three main types exist:
- the basic epidermis cell
- the cells of the stoma complexes and
- the trichomes (gr.: trichoma = hair), epidermal attachments of varying shape, structure and function
The basic epidermis cells, i.e. the least specialized cells constitute the largest group of dermal cells. They seem either polygonal or elongated in top view. Their walls are often wavy or sinuate. It is unknown, what induces this shape during development, since the explanations given by the existing hypotheses seem insufficient. Elongated epidermis cells can be found at organs or parts of organs that are elongated themselves, like stems, leaf petioles, leaf veins or leaves of monocots. The epidermises of the leaf's upper- and undersurface may have different structures. The shape of the cells, the thickness of the walls as well as the distribution and number of specialized cells (guard cells and trichomes) per area may all vary. Wide varieties of different cell shapes may even exist in species of a single family, e. g. in the Crassulaceae family
The wall of epidermal cells that constitute the leaf's surface is often thicker than the other walls. This can be particularly well observed with the epidermis of conifer needles and that of xerophytes (plants living in dry habitats). Aquatic plants have usually thin walls. The wall of many seeds becomes stronger during ripening and may fill nearly all of the cell's lumen so that the protoplast is driven out and degenerates. The basic epidermal cells of most species contain no chloroplasts. Some ferns and several aquatic or shade plants are exceptions.
The epidermis is more often than not built from a
single cell layer, though multi-layered,
water-storing epidermises that evolved from initially single-layered
tissues by periclinal division have been shown among the species of
several families (Moraceae: most Ficus-species,
Piperaceae: Peperonia,
Begoniaceae, Malvaceae and others).
Epidermis cells secrete a cuticle, that covers
all epidermal surfaces like an uninterrupted film. It may either be
smooth or structured by bulges, rods, filaments, folds,
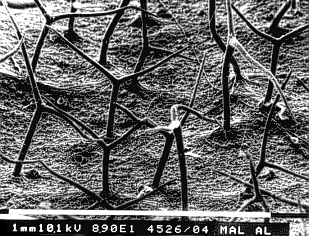
or furrows. 
The folding of the cuticle is nevertheless not always caused by cuticular rods or filaments. There are cases where it is merely the expression of the bulging of the cell walls below. It may be necessary to analyze cross-sections of the cuticle and the underlying epidermal cells to find the actual cause.
In some cases, like that of the tomato fruit, the cuticle is pigmented with carotinoids. Often, additional waxes, oils, resins, salt crystals and (hydrophilic) mucilage are excreted. The latter is especially common in developing seeds. The excretion of wax inhibits the moistening of the leaves more than the cuticle itself. These waxes do often have an intrinsic structuring. A thick layer of wax lends a whitish appearance to the plant surface. It has two effects: diminution of water-loss and enhancement of reflection of the sunlight, thus protecting the plant from too much heat.
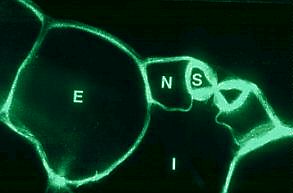
Cross-Section Through a Stomate of Rhoeo discolor . S Guard cell, N Subsidiary cell, E Non-specialized epidermis cell, I Intercellular space. (R. KAPPLER, 1984)
The stomata accomplish an important function of the epidermis. Their
complete functional unit is the stoma
complex. It is composed of two chloroplast-containing
guard cells that enclose a pore
(the actual stoma), as well as of two to four
subsidiary (or accessory)
cells devoid of chloroplasts.
Cross-sections show that the guard cells have walls of different
thicknesses. They sit at the top of an
intercellular space, that is in
communication with other intercellular spaces of the respective
tissue. The stoma can be opened or closed according to demand. In
this way, the guard cells regulate both transpiration and
CO2- uptake. Water-content and CO2-
concentration within the plant tissue act as regulators for the state
of opening. The guard cells control the size of the stoma by changing
its shape: more about the mechanism.
Stomata occur at all plant surfaces above ground. Their number approaches 100-300 per mm2, but there exists a wide variation in number and distribution. They are lacking in roots, in the epidermis of the chlorophyll-free shoot of the parasitic species Monotropa hypopitys and Neottia nidus-avis as well as in some submerse living aquatic plants, while they are normal in others. They do, too, happen to be present in some colored or white petals, though they have often lost their function here. The parallely veined leaves of most monocots, some dicots and the needles of conifers have stomata that are grouped in parallel rows.
Stomata are produced by the guard cell mother cells, that again are laid out within the epidermis in regular intervals. The initial of the stoma complex is the smaller of the two progenitor cells. It contains more plasma. Both guard cells stem from it and are formed by unequal division, while the production of the subsidiary or accessory cells may occur in many different ways. The development of a leaf's stoma happens mostly asynchronously. It follows the successive differentiation of the single tissues in leaves with parallel venation. Differentiation proceeds in a basipetal manner, i.e. it starts at the tip of the leaf and continues towards its base.
Netted leaves have stomata of different developmental stages
spread mosaic-like across the whole lamina.
Variations. Morphological variations of stoma complexes can be expected, since different ways of their development exist. The guard cells of gramineous plants are a typical exception. They are often shaped like dumb-bells.
 The
hydathodes, that can often be
found at the end of vascular bundles are other derivatives of stoma
complexes. Their guard cells do still look like those of normal stoma
complexes, but they cannot be closed any more. Water secretion by the
hydathodes is called guttation.
Characteristic hydathodes occur especially at the leaf margins of
Garden Nasturtium (Tropaeolum
majus), Lady's Mantle (Alchemilla vulgaris - picture to
the right: from J. v. SACHS, 1887) and at the leaves tips of many
grasses. Salt, sugar and organic compounds that are dissolved in the
guttation water crysalize after evaporation of the water at the site
of outflow. Typical examples are the lime secretions of
Saxifrage-species (Saxifraga) and the
salt glands of halophytes.
The
hydathodes, that can often be
found at the end of vascular bundles are other derivatives of stoma
complexes. Their guard cells do still look like those of normal stoma
complexes, but they cannot be closed any more. Water secretion by the
hydathodes is called guttation.
Characteristic hydathodes occur especially at the leaf margins of
Garden Nasturtium (Tropaeolum
majus), Lady's Mantle (Alchemilla vulgaris - picture to
the right: from J. v. SACHS, 1887) and at the leaves tips of many
grasses. Salt, sugar and organic compounds that are dissolved in the
guttation water crysalize after evaporation of the water at the site
of outflow. Typical examples are the lime secretions of
Saxifrage-species (Saxifraga) and the
salt glands of halophytes.
Epidermal attachments of various shape, structure and function are called trichomes. They protect and support the leaf, produce glands in the form of scales, different papills and, in roots, often absorbing hairs. They stem exclusively from epidermal cells. Often a trichome is formed by just one cell though sometimes several cells are involved.
Trichomes have to be distinguished from
multi-cellular trichomes (like
spines) and from short shoots (like
thorns), since these do contain not only cells of epidermal origin
but of other tissues, too.
Hairs can be found at plant
surfaces in manifold shapes. They can be single- or multi-celled,
branched or unbranched, living or dead. Their walls may be
strengthened by silicate, calcium carbonate or other encrustations
giving the respective hairs a bristle-like appearance. Such stiff
bristles (at Boraginaceae and Cruciferaceae for example) protect
plants from being eaten away by animals. Many hairs, particularly the
strongly branched ones, help avoiding transpiration losses. It is
well known that plants from dry habitats are either succulent (and
thus have a thick cuticle) or densely covered with silvery hair. 120
hairs per mm2 were counted at Stachys lanata
(woundwort). Their microscopic analysis shows strongly branched and
dead hair cells. These hairs offer three advantages:
- The lumen of dead cells is air-filled. It gives them a silvery-whitish appearance. A large proportion of the incoming light is thus reflected. A thick layer of wax has the same effect.
- Only weak circulation takes place at the leaf surface, thus reducing water loss to a minimum.
- Dying of the hair cells reduces the surface area, where water could be lost drastically. If the cells were living, too a high loss of water loss would occur due to the largely increased surface of the branched hair cells.
Hairs are organized in a regular pattern on the leaf surface. They
are laid out at nearly constant distances. Comparable patterns can be
found in the lay out of stomata. Why this regularity? No definite
answer can be given until today, but we can present a mathematical
model that follows rules, that may very likely be valid for plant
tissues, too.

Aerenchyma with lignified "inner hairs" (Nymphaea spec.): section through the leaf stalk. Staining with phloroglucin-HCl. (photo by W. KASPRIK).
A kaleidoscope of different hairs. Exceptionally long (1-6 cm), single-celled and unbranched hairs out of nearly pure cellulose wrap up the seeds of Gossypium (cotton). With Lobelia, Arabis alpina or Malacantha alnifolia (see picture to the right), they are branched. Multi-celled hairs can consist of one or several rows of cells. Typical examples are the dendritic trichomes of the plane tree (Platanus hybrida) or those of the dark mullein (Verbaseum nigrum). Hairs at the leaves of oaks (Quercus robur) look like tufts that of many Malvaceae are stellate. Those of the oleaster (Elaeagnus angustifolia) are formed like scales. Bromeliaceae use such scales for water absorption. The humidity of the air is collected within the scales with the help of capillary forces and later on used by the plant. Gland hairs consist of a single-celled stalk and a single- or multi-celled head.
Some examples include:
The gland hairs at the leaves of tobacco (Nicotiana tabacum) and e. g. Adenocaulon (Compositae) have multi-celled heads, while they are single-celled with primroses (Primula sinensis) and garden geraniums (Pelargonium zonale,for example). The secretion of Pelargonium zonale and Origanum (Lamiaceae) is a volatile oil. The gland cells themselves are rich in plasma, secretion takes place through the cell walls. The oil gathers at the cell surface and can be seen as a cap that refracts light strongly and is covered by a thin film of both cuticle and cell wall components. After the cap's splitting-off, the oil is set free. The peel of citrus fruits contains another type of secretory glands that is sunken into the epidermis.
The filaments of Tradescantia virginiana are surrounded by multi-celled hairs. The single cell looks like a barrel. The protoplast lives at the wall and the vacuole is colored with a purple dye from the anthocyane-family. It is crossed by numerous plasma chords. The nucleus is located in the cell's center looking like it were strung up on the plasma chords. Since the plasma chords keep changing their shape all the time, the nucleus is torn to and fro as if at rubber bands. The plasma of the chords has a strong and lively current and transports plainly visible granules. The current is strictly directed and currents of opposing directions can often be seen in other chords. The hairs on the filaments of Tradescantia virginiana are a classic example for currents, since they are easily perceived and very regular in organization. Consult this site about the molecular mechanisms of plasma currents.
The hairs at the filaments of the hardheads (Centaurea
jacea) and those of the cornflower (Centaurea cyanus) got
well-known for other reasons. They are sensitive to touch and cause
the filaments to move.
The stinging
hairs of the stinging nettle are really multi-cellular
trichomes. They consist of two parts and a multi-celled base that
develops not only from epidermal cells, but from those of the
subepidermal layers, too. A hair cell is sunken into the base. Its
basal part is called bulbus. It is surrounded by the cells of the
base like liquid is surrounded by a mug. Its upper part is elongated
and thin and ends in a laterally attached head. At the site of
transition, the cell wall is considerably thinner than in other parts
of the cell. Encrusting silicates make it brittle and cause the head
to break off easily when touched. This leaves the hair with a point
that bears an astonishing resemblance to the needle of a syringe. The
pressure of the touch is directly transferred to the bulbus due to
the rigidity of the cell wall. It presses the content of the bulbus
(sodium formamide, acetyl choline, histamine) through the cannula and
injects it into the wound.
Further Types of Trichomes And Special Functions of The Epidermis.
Papillae are protrusions of the epidermal surface. Typical textbook-examples are the papillae of the pansy flower (Viola tricolor) as well as the leaf surfaces of many species from the rain forest. They give a velvet appearance to the plant's surface. Some epidermal cells may be developed into water-storing vesicles.
The water vesicles at the surfaces of many Mesembryanthemum-species and other succulent species are such derivatives. With some plants, like bellflower (Campanula persicifolia), the outer walls of the epidermis are thickened like a lens. They collect the light, which is then perceived by specialized light receptors (also called perceptors) and used for physiological reactions.

Bladder cell in top view. Epidermal imprint. Monanthes lowei. (preparation J. THIEDE)