Botanik online 1996-2004. Die Seiten werden nicht mehr bearbeitet, sie bleiben als historisches Dokument der botanischen Wissenschaft online erhalten!
Proteine als analytische Hilfsmittel (Sonden) zur Lokalisation
von Molekülen und zum Studium der molekularen Architektur
der Zellen
Seit der zweiten Hälfte des vorigen Jahrhunderts werden in
der Mikroskopie Farbstoffe zum Nachweis bestimmter Stoffklassen
verwendet. In den dreißiger Jahren dieses Jahrhunderts kamen
Fluoreszenzfarbstoffe hinzu, die - in wesentlich niedrigeren Konzentrationen
einsetzbar - auch als "Vitalfarbstoffe" Verwendung fanden.
Farbstoffmoleküle sind meist klein, und ihr Nachteil ist
daher eine nur begrenzte Spezifität. Es gibt u.a. proteinspezifische
Farbstoffe, doch kaum einen, der für ein bestimmtes Enzym
spezifisch wäre. Die selektive Bindung wurde bei einigen
Toxinen (z.B. Pilzgiften) nachgewiesen. Das Phalloidin (Toxin
des Weißen Knollenblätterpilzes) bindet an Aktin. Durch
Kopplung eines Fluoreszenzfarbstoffs an das Phalloidin läßt
sich demnach Aktin in der Zelle lokalisieren.
Weit spezifischer sind die verschiedenen Makromoleküle und
hier vor allem die Proteine:
Lektine: Im vorletzten Abschnitt haben wir die Lektine
kennengelernt. Jedes kann mit einem Fluoreszenzmarker,
wie Fluoroisothiocyanat (FITC: grüne Fluoreszenz) oder Rhodamin
(rote Fluoreszenz) versehen werden. Solche Präparate werden
in der medizinischen Forschung in großem Stil eingesetzt
und sind daher relativ preiswert im Handel erhältlich. Sie
eignen sich als Sonden zur Lokalisierung von Glykokonjugaten (=Lektinrezeptoren)
an Zelloberflächen, an Membranoberflächen, in Kompartimenten
u.a.
Makromoleküle haben den Nachteil, nicht ohne weiteres in
Zellen eindringen zu können. Sie sind daher primär Marker
extrazellulärer Oberflächenrezeptoren. Durch Cellulasebehandlung
z.B. kann die Wand pflanzlicher Zellen abgebaut werden; es entstehen
zellwandlose Protoplasten. Je nach Herkunft binden sie ConA oder
RCA, was darauf hinweist, daß Glucose- oder Mannosereste,
respektive Galactosereste an ihrer Oberfläche exponiert sind.
Protoplastenpräparationen enthalten vielfach die unterschiedlichsten
Zellfragmente, unter anderem auch freie Vakuolen. Die Vakuolenmembran,
der Tonoplast, bindet keines der bekannten Lektine. Allein schon
damit ist gezeigt, daß er chemisch anders aufgebaut ist
als die Plasmamembran.
Fluoreszenzmarkierte Lektine eignen sich unter anderem zur Kartierung
von Lektinrezeptoren in oder an Zellwänden, zum Nachweis
einer Polarität von Zellen, zum Nachweis unterschiedlicher
Aktivitätszustände oder bestimmter Stadien des Zellzyklus;
sie eignen sich zur Messung der Löslichkeitseigenschaften
der Lektinrezeptoren. WGA ist ein Indikator für Pilzmycelien
in infiziertem pflanzlichem Gewebe.
Für elektronenmikroskopische Studien müssen die Lektine
mit elektronendichten Markern (z.B. Ferritin oder kolloidalem
Gold) gekoppelt werden. Mit diesen Komplexen lassen sich in entsprechend
vorbereiteten Präparaten u.a. unterschiedliche Verteilungen
an Membraninnen- und außenseiten erkennen. Lektine haben
natürlich auch Nachteile:
- Ihre Bindungsorte sind die sogenannten Glykokonjugate. Es
ist daher ohne Zusatzanalyse nicht möglich, zu entscheiden,
ob z.B. ein ConA-Rezeptor ein Oligo- oder Polysaccharid, ein Glykoprotein
oder ein Glykolipid ist.
- Lektine sind nur gegen ein sehr enges Spektrum an Zuckern
gerichtet. Es gibt keine Lektine, um z.B. Arabinose- oder Xylosereste
zu lokalisieren.
 Antikörper sind eine recht homogene Gruppe von Proteinen,
die im Serum von Vertebraten vorkommen und die den tierischen
Organismus vor Fremdeinflüssen, beispielsweise vor Infektionen
mit Bakterien und Viren oder vor eigenen Tumorzellen schützen
sollen. In Pflanzen kommen keine Antikörper vor. Da wir sie
aber zum Studium von Pflanzenzellen brauchen, hier zunächst
ein kurzes Porträt:
Antikörper sind eine recht homogene Gruppe von Proteinen,
die im Serum von Vertebraten vorkommen und die den tierischen
Organismus vor Fremdeinflüssen, beispielsweise vor Infektionen
mit Bakterien und Viren oder vor eigenen Tumorzellen schützen
sollen. In Pflanzen kommen keine Antikörper vor. Da wir sie
aber zum Studium von Pflanzenzellen brauchen, hier zunächst
ein kurzes Porträt:
Die Antikörperbildung ist induzierbar (Immunisierung). Das
heißt, es wird ein Signal benötigt, das den tierischen
Organismus zur Antikörperbildung stimuliert. Ein solches
Signal muß stets makromolekular sein; es kann ein Bestandteil
einer Zelloberfläche sein. Man bezeichnet solche Komponenten
als Immunogene oder Antigene.
Ein Antigen ist meist größer als die Bindungsstelle
eines Antikörpers. Es kann daher mehrere (verschiedene) Antikörpermoleküle
binden. Der antikörperbindende Bereich des Antigens ist eine
antigene Determinante. Bei einer Immunisierung werden so viele
verschiedene Antikörper gebildet, wie es antigene Determinanten
gibt. Die Antikörperpopulation ist daher stets heterogen
oder, wie man heute sagt, polyklonal.

Antigen-Antikörperreaktion.
Antikörper (Y-förmig dargestellt) stellen eine heterogene
Molekülpopulation mit unterschiedlichen Spezifitäten
dar. Eine Kreuzreaktion einer Antikörperpopulation (eines
Antiserums) mit einem Fremdantigen (Mitte) erfolgt nur dann, wenn
das homologe und das Fremdantigen zumindest partiell mit den gleichen
Determinanten bestückt ist. Jedes Antikörpermolekül
besitzt zwei gleiche Bindungsstellen für antigene Determinanten.
Um Antikörper mit einer bestimmten Spezifität zu erzeugen,
immunisiert man üblicherweise Kaninchen. Zwei bis drei Wochen
nach der Immunisierung kann ihnen Blut entnommen werden. Nach
dem Abzentrifugieren der Blutzellen erhält man das sogenannte
Antiserum mit den darin enthaltenen spezifischen Antikörpern.
Sie eignen sich nunmehr zum qualitativen und quantitativen Nachweis
des eingesetzten Antigens sowie zum Nachweis von Substanzen, die
diesem Antigen ähnlich sind. Man spricht dabei von serologischer
Kreuzreaktion oder serologischer Verwandtschaft. So findet man
in der Regel eine mehr oder weniger stark ausgeprägte serologische
Verwandtschaft zwischen homologen Proteinen (Enzymen, Speicherproteinen,
dem Cytochrom c usw.), die man aus mehr oder weniger nah verwandten
Tier- oder Pflanzenarten gewonnen hat. Der serologische Verwandtschaftsgrad
ist hier meist mit der Zahl der Aminosäureunterschiede zwischen
den Proteinen korreliert.
Es gibt eine Anzahl zum Teil sehr empfindlicher serologischer
Nachweisverfahren, z.B. den Radioimmuntest
(radio immuno assay: RIA).
Benötigt man einen Antikörper gegen ein kleikes Molekül,
z.B. gegen ein Phytohormon, so muß man es als erstes an
einen makromolekularen Träger koppeln; dadurch nimmt es die
Eigenschaft einer antigenen Determinante an, und unter den zahlreichen,
gegen diesen Komplex gebildeten Antikörpern sind auch solche,
die gegen das Phytohormon gerichtet sind. Sie sind von den übrigen
leicht abtrennbar, denn alle anderen reagieren mit dem Träger
allein und können daher durch Präzipitation aus dem
Serum entfernt werden. Übrig bleiben dann nur noch die hormonspezifischen
Antikörper.
Zur Lokalisation von Antigenen in Zellen geht man den schon für
Lektine beschriebenen Weg. Es ist jedoch nicht üblich, den
Antikörper direkt mit einem Fluoreszenzmarker zu versehen,
denn es ist viel vorteilhafter, in einem anderen Tier (z.B. in
Ziegen) Antikörper gegen Kaninchen-Antikörper zu erzeugen,
diese dann in großen Mengen zu gewinnen und sie mit dem
Fluoreszenzfarbstoff zu markieren. Man spricht hier von indirekter
Immunfluoreszenz. Warum dieser Umweg?
Ziegen-anti-Kaninchen-Antikörper können gegen jeden
beliebigen in Kaninchen erzeugten Antikörper verwendet werden.
Von den spezifischen Kaninchen-Antikörpern stehen meist
nur geringe Mengen zur Verfügung. Vielfach werden parallel
eine Anzahl von Kaninchen mit unterschiedlichen Antigenen immunisiert.
Man müßte demnach jeden der produzierten Antikörper
getrennt mit dem Fluoreszenzfarbstoff markieren.
Man erhält eine Amplifikation, eine Verstärkung
der Fluoreszenz, weil mehrere Ziegen-Antikörper von einem
Kaninchen-Antikörpermolekül gebunden werden.
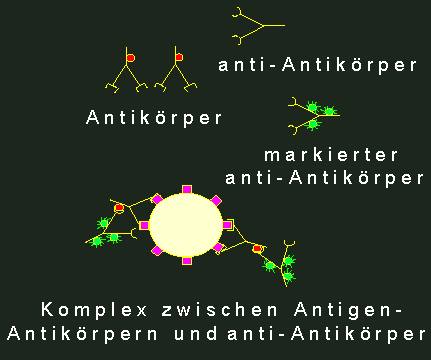
Indirekte
Immunfluoreszenz. l. Spezifische Antikörper gegen das primäre
Antigen. Die Antikörper selbst tragen antigene Determinanten
(durch rote Kreise markiert). Normalerweise stellt man Antikörper
dieser Art in Kaninchen her. 2. anti-Antikörper (hergestellt
z.B. durch Immunisierung einer Ziege mit Kaninchenantikörpern).
Diese anti-Antikörper (Ziege gegen Kaninchen) binden an antigene
Determinanten des Kaninchenantikörpers. Wenn man sie vorher
mit einem Fluoreszenzmarker (grün) versehen hat (3),
erhält man am primären Antigen einen fluoreszierenden
Komplex (4).
Für elektronenmikroskopische Untersuchungen verwendet man
auch hier wieder elektronendichte Marker, mit denen die Ziegen-anti-Kaninchen-Antikörper
versehen werden.
So umfangreich der Einsatz von Antikörpern in der medizinischen
Forschung auch ist, so spärlich sind die Anwendungen bisher
zum Studium von Pflanzenzellen. Einer der Gründe dafür
ist die Zellwand, die von keinem Antikörper durchdrungen
wird, zum anderen der Mangel an geeigneten Antigenen. Dennoch
gibt es vorzeigbare Ergebnisse. Das Phytochrom, ein Rezeptor für
Licht, das zu den wichtigsten Kontrolleinheiten in Pflanzenzellen
gehört, konnte in bestimmten Zellen lokalisiert
werden; es konnte aber auch gezeigt werden, daß es in anderen
fehlt.
Zur Markierung von Zellinhalten müssen die Zellen entweder
angeschnitten werden (z. B. Herstellung von Gefrierschnitten),
oder es müssen Protoplasten erzeugt werden, deren Membran
partiell durchlässig ist. Zu den weiteren durch Immunfluoreszenz
lokalisierten Antigenen gehören u.a. einige Enzyme wie die
Phosphoenolpyruvatcarboxylase, die alpha-Amylase sowie
einige der Speicherproteine und der Elemente des Cytoskeletts.
Seit der Entdeckung durch C. MILSTEIN und G. KÖHLER (Medical
Research Council, Laboratory of Molecular Biology, Cambridge und
Basel Institute of Immunology) im Jahre 1975, werden die sogenannten
monoklonalen Antikörper als Non plus ultra gehandhabt.
Es sind homogene Antikörperpopulationen, die in Zellkultur
produziert werden und die ein sehr enges Spezifitätsspektrum
aufweisen (nur gegen eine Determinante gerichtet). Zu ihrer Herstellung:
Es muß vorab noch gesagt werden, daß Antikörper
in kleinen Lymphozyten produziert werden. Eines der lymphozytenreichsten
Gewebe ist die Milz. Man immunisiert nunmehr - wie gehabt - z.B.
eine Maus und entfernt ihr nach einigen Tagen die Milz, zerkleinert
sie und gewinnt somit eine Zellsuspension. Diese Zellen werden
mit Myelomzellen der Maus (einer von vielen Tumorzellinien) fusioniert.
Durch die Fusion entsteht eine Hybridzelle (ein Hybridom). Myelomzellen
zeichnen sich durch uneingeschränktes Wachstum aus; diese
Eigenschaft bleibt auch in den Hybridomzellen erhalten. Hinzu
kommt deren Eigenart, Antikörper zu bilden und zu sezernieren.
Es bedarf eines zwischengeschalteten Selektionsschrittes, um eine
gewünschte Zellinie zu isolieren und sie dann in Kultur zu
nehmen.
Der Nachteil des Verfahrens: Die Antikörper sind zwar hochspezifisch,
aber die Zahl der Bindungsstellen (Antigene) in den Zellen ist
entsprechend verringert. Es werden daher nur sehr wenige Antikörpermoleküle
gebunden, oft liegt die Antigen-Antikörper-Reaktion deshalb
an der Grenze der Nachweisbarkeit. Aber auch hier sind Wege vorgezeichnet,
um zum Erfolg zu gelangen: Im Falle der indirekten Immunfluoreszenz
bedient man sich TV-Kameras mit elektronischer Restlichtverstärkung.
Lokalisierung von Enzymaktivitäten. Lassen sich Enzyme
oder Substrate in der Zelle auch ohne den Aufwand mit den Antikörpern
lokalisieren?
Auch hier gibt es verschiedene Ansätze. Am gebräuchlichsten
ist die Autoradiographie. Man benötigt ein radioaktiv markiertes
Substrat, besser ein Substratanaloges, das zwar vom Enzym gebunden,
von ihm aber nicht umgesetzt wird. Nach dem Auswaschen von nichtgebundenem
Material kann durch Auftragen eines Spezialfilms auf die Zelle
ein Autoradiogramm hergestellt werden, aus dem die Position der
Enzyme ersichtlich ist. Das Verfahren wird vornehmlich bei membrangebundenen
Enzymen eingesetzt.
Eine andere Möglichkeit ist die Verwendung eines Substrates,
das umgesetzt wird und als Produkt ein - möglichst unlösliches
- farbiges Produkt ergibt; für die Elektronenmikroskopie
verwendet man auch hier wieder elektronendichte Marker.
© Peter v. Sengbusch - Impressum