Botanik online 1996-2004. Die Seiten werden nicht mehr bearbeitet, sie bleiben als historisches Dokument der botanischen Wissenschaft online erhalten!
Die bereits besprochenen Ergebnisse von W. BATESON u.a., daß bei Lathyrus odoratus zwei Merkmale in der Regel gekoppelt sind, gelegentlich aber voneinander getrennt werden, bilden einen experimentellen Ansatz zur Lokalisierung von Genen auf den Chromosomen. BATESONs Daten waren noch nicht ausreichend, um dementsprechende Konsequenzen ziehen zu können. Es waren schließlich wiederum die an Drosophila gewonnenen Ergebnisse, die diesen Schluß zuließen.
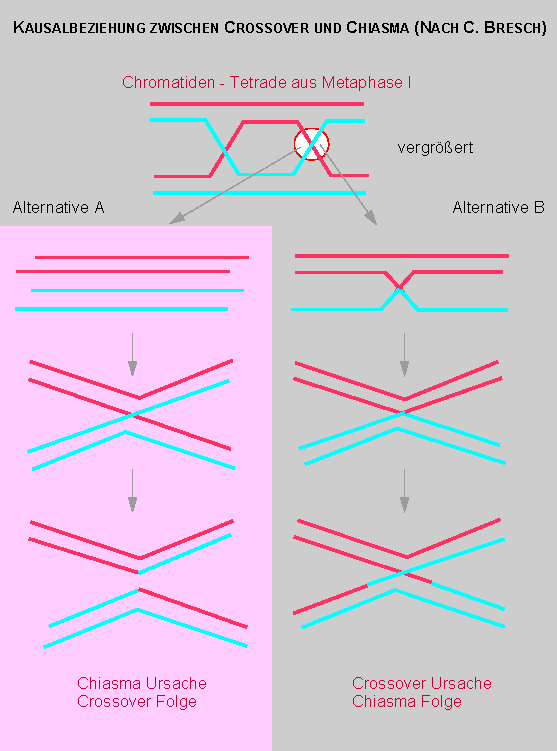
Die Chromosomentheorie (in ihrer einfachsten Formulierung) fordert, daß die Zahl der Kopplungsgruppen der Zahl der Chromosomen (1n = haploider Satz) entspricht. Tatsächlich wurden aber, zunächst bei Pisum sativum, später auch bei anderen Arten, mehr Kopplungsgruppen als 1n Chromosomen nachgewiesen. H. de VRIES vermutete daher (1903), daß die Vermehrung der Zahl der Kopplungsgruppen auf Chromosomenstückaustausch beruht. Die strukturellen Voraussetzungen dafür sind während der Prophase der Meiose gegeben, in der sich homologe Chromosomen paaren. F. A. JANSSENS beobachtete während der Meiose von Batrachoseps attenuatus (einem Amphibium) die Überkreuzung einzelner Chromosomenarme im Diplotänstadium und nannte diesen Zustand Chiasma. Er erkannte darüber hinaus, daß die Chromosomenarme (genauer Chromatidenarme) an dieser Stelle brechen können und über Kreuz wieder verwachsen.
T. H. MORGAN zeigte darauf an zwei X-chromosomengebundenen Merkmalen, daß ein Faktorenaustausch erfolgt und daß dieser auf einem Crossing-over beruht. Mit anderen Worten: aufgrund genetischer Analysen wird ein Chromosomenstückaustausch gefordert.
Bei der Aufstellung einer Genkarte ist die relative Crossing-over-Wahrscheinlichkeit zwischen zwei Genen (Genorten) entscheidend. Liegen zwei Genorte auf einem Chromosom nahe beieinander, ist die Wahrscheinlichkeit ihrer Trennung durch Crossing-over gering; mit steigendem Abstand nimmt sie zu. Unter dieser Prämisse wertete MORGANs Mitarbeiter A. H. STURTEVANT die Austauschhäufigkeiten mehrerer auf dem X-Chromosom liegender Gene aus. Er zeigte damit, daß sich die Austauschhäufigkeiten mit den Genabständen korrelieren ließen und daß sich Genabstände additiv verhielten. So ist beispielsweise der Abstand von A-C gleich der Summe aus A-B + B-C. Dies wiederum heißt, daß die Genorte A, B und C linear auf dem Chromosom angeordnet sind. Die Werte für große Genabstände sind jedoch oft niedriger als erwartet (niedriger als die Summe kurzer Teilstrecken). Dieses Phänomen, Interferenz genannt, beruht auf der Beeinflussung der Wahrscheinlichkeit des Auftretens eines weiteren Crossing-over Ereignisses durch ein vorangegangenes.
Der ersten, anfangs noch sehr einfachen Genkarte einer Kopplungsgruppe (X-Chromosom von Drosophila) folgten bald weitere sowie Ergänzungen der ersten. Bereits 1926 lag auch die erste Genkarte (mit nur drei Genorten) eines pflanzlichen Chromosoms vor (L. J. STADLER, University of Missouri, Columbia, 1926).
Neben dem meiotischen Crossing over ist mitotisches Crossing over mit Schwesterchromatidaustausch bekannt..
Nach der Kartierung zahlreicher neuer Genorte mußten die ursprünglich von STADLER festgelegten Abstände präzisiert werden, da auch sie durch Doppel-Crossing-over verfälscht waren. Erst die Feinanalyse unter Zuhilfenahme zahlreicher Genorte (Marker) führte in Annäherung zu den realen Abständen.