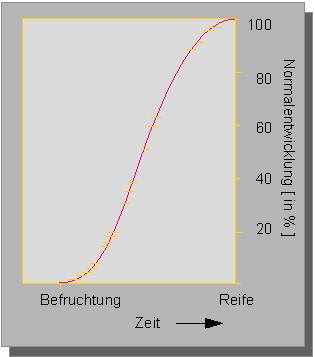
Botanik online 1996-2004. Die Seiten werden nicht mehr bearbeitet, sie bleiben als historisches Dokument der botanischen Wissenschaft online erhalten!
C. DARWIN (1868) hat sich über einen langen Zeitraum hinweg mit botanischen Problemen befaßt, arbeitete über Selbst- und Fremdbefruchtung, hat verschiedene Arten miteinander gekreuzt und die Nachkommenschaft zahlenmäßig erfaßt, ohne irgendwelche Schlüsse aus den Zahlen zu ziehen. Unter anderem arbeitete er mit dem Löwenmäulchen (Antirrhinum majus), einer Art aus der Familie der Scrophulariaceae. Die Blüten der meisten Arten dieser Familie sind zygomorph, also nur durch eine Symmetrieebene gekennzeichnet. Wenige Arten sind (wie die meisten Blüten der Angiospermen) radiärsymmetrisch. Gelegentlich tritt dieser Blütentyp auch bei Antirrhinum majus und anderen Arten (z. B. Digitalis purpuraea) auf. Nach Isolation und Selbstung (Bestäubung mit Pollen des gleichen Individuums) kann eine reine Linie etabliert werden. DARWIN kreuzte derartige Pflanzen mit zygomorph blühenden Partnern und erhielt folgendes Ergebnis:
Zygomorphie und Radiärsymmetrie bei Antirrhinum majus |
|||
| Parentalgeneration: |
|
||
F1 : |
|
||
|
F2 : |
zygomorph | intermediär | radiärsymmetrisch |
|
|
|
|
|
Schlägt man die intermediären Formen den zygomorphen zu, ist das Ergebnis mit einer 3:1-Spaltung vereinbar. Zygomorph ist offensichtlich ein dominantes Merkmal, wobei die Dominanz allerdings unvollständig ist, denn es gibt eine ganz schwache Tendenz in Richtung intermediär.
C. CORRENS (seinerzeit am Botanischen Institut der Universität Leipzig) analysierte eine große Zahl von Arten und Merkmalen und suchte Antworten auf verschiedenste Fragen. In einer 1902 erschienenen Arbeit beschrieb er eine Spaltung beim Mais (Zea mays):
rundliche Körner (dominantes Merkmal) : runzlige Körner (rezessives Merkmal):
in der F2: 8975 : 1711.
Bei einer 3 : 1-Spaltung wären die Werte 8015 : 2671 zu erwarten. Die Abweichung schien ihm zu groß zu sein, und er erklärte sie durch ein mehr oder weniger leichtes Zustandekommen der einen oder der anderen Keimzellkombination. Diese Aussage ist insofern wichtig, als die MENDELschen Regeln auf statistischen Annahmen beruhen und u.a. davon ausgehen, daß alle Gametentypen stets in gleicher Menge und alle Kombinationen mit gleicher Wahrscheinlichkeit auftreten. Lebende Systeme sind aber, wie auch die nachfolgenden Beispiele zeigen werden, keineswegs immer als gleichwertig anzusehen. DARWINs Selektionstheorie der Evolution baut bekanntlich auf der Existenz ungleicher Genotypen in einer Population auf.
Ein weiteres, von CORRENS untersuchtes Versuchsobjekt ist die Brennessel. Die Art Urtica pilulifera hat Blätter mit gesägtem, Urtica dodartii Blätter mit glattem Rand. Gesägt ist dominant über glatt. In der F2 aus einer Kreuzung beider Arten kommt es daher zu einer klaren 3:1-Spaltung. Dieses Beispiel ist in zahlreichen Lehrbüchern zitiert, das Schema abgebildet worden. Bei Pflanzen mit gesägten Blättern gleichen die homozygoten Formen augenscheinlich den heterozygoten. Bei den Heterozygoten zeichnen sich jedoch die beiden ersten Laubblätter (und nur diese) durch eine geringere Zahl an Blattrandzähnchen aus, sie sind dadurch von den entsprechenden Laubblättern der Homozygoten unterscheidbar. CORRENS nannte diese Erscheinung eine versteckte Hinneigung zur intermediären Form.
Bei dem klassischen MENDELschen Erbsenbeispiel (Samenform rund:kantig) bezieht sich die Dominanz, wie wir ja schon gesehen haben, auf die Samenform. Das gleiche Gen beeinflußt aber auch die Form der Stärkekörner. In runden Samen findet man einfach gebaute, große, längliche, in runzligen Samen eigenartig zusammengesetzte Stärkekörner. In den Heterozygoten (in der Fl und F2) kommen beide Formen nebeneinander vor (A. D. DARBISHIRE, 1911); sie stellen damit einen intermediären Typus dar. Die Erscheinung, daß ein Gen mehrere Merkmale beeinflußt, heißt Pleiotropie.
Es wird oft unterschätzt, wie breit die wissenschaftlichen Kenntnisse jener waren, die die Genetik von Anbeginn gestaltend und richtunggebend beeinflußt haben. Vor und nach der Jahrhundertwende wurde eine Fülle von Bastardierungen durchgeführt. Doch nur ganz wenige Beispiele gingen in die Lehrbuchliteratur ein, und es mag daher der falsche Eindruck aufkommen, nur sie seien angesetzt und ausgewertet worden. Alles, was an pflanzlichen Objekten gezeigt werden konnte, ließ sich auch bei Tieren verifizieren. Die immer wieder gebrachten Beispiele geben die Verhältnisse am anschaulichsten wieder und demonstrieren zum Teil wenigstens eine Reihe scheinbarer Abweichungen.
Hinzu kommt, daß der Begriff "Merkmal" nicht klar definiert ist. Es hängt allein vom Betrachter ab, was er darunter versteht, und nicht immer sind die Alternativen so klar wie bei dem Merkmalspaar Blütenfarbe rot/weiß. Nun weiß man aber auch, daß es bei vielen Blütenfarben Farbschattierungen und zahlreiche Übergänge gibt, und für alle diese Erscheinungen müssen schließlich Erklärungen gefunden werden. Landwirte und Pflanzenzüchter sind in erster Linie am "Ertrag" interessiert, einem Merkmal, von dem jeder Praktiker weiß, daß es einem hohen Grad an Variabilität unterliegt.
Die Definition von "Merkmal" beschränkt sich keineswegs auf ohne Hilfsmittel erkennbare Phänotypen. Das Mikroskop, andere physikalische und (bio)- chemische Analyseverfahren wurden zu genauso wichtigen Hilfsmitteln der Genetiker wie effiziente Auslese (= Selektions)- verfahren und eine mathematische Auswertung der Ergebnisse. Je mehr man sich mit der Problematik, wie ein Gen wirkt oder was ein Gen ist, befaßte, um so mehr mußte die Methodik verfeinert werden, um so wichtiger wurde es, ein geeignetes (und möglichst einfaches) Objekt zu analysieren, und um so mehr Vorkenntnisse waren erforderlich, um einen komplexen Vorgang Schritt für Schritt zu entschlüsseln.
Wieder eine Erscheinung, die auch C. DARWIN untersucht hatte. Viele Primelarten zeichnen sich durch Heterostylie aus. Etwa die Hälfte der in der Natur vorhandenen Blüten enthalten einen langen, die anderen einen kurzen Griffel. Die Staubbeutel befinden sich niemals auf Griffelhöhe. Primeln sind durchweg Fremdbefruchter, und normalerweise sind langgrifflige Formen nur durch Pollen kurzgriffliger, und kurzgrifflige nur durch Pollen langgriffliger befruchtbar, womit ein wirkungsvoller Schutz vor Selbstbefruchtung gegeben ist. Bei einigen wenigen Arten (z.B. Primula sinensis) kann (unter Ausschluß fremden Pollens) eine Selbstbefruchtung ausgelöst werde. Bei Selbstung langgriffliger Formen erhält man in der folgenden Generation ausschließlich langgrifflige Blüten, bei einer Selbstung kurzgriffliger entstehen kurzgrifflige und langgrifflige Blüten im Verhältnis 3:1, und bei einer Kreuzung langgrifflig x kurzgrifflig (wie in der Natur) erscheinen beide Formen im Verhältnis 1:1.
Die Befunde lassen sich einfach dadurch erklären, daß Kurzgriffligkeit auf Heterozygotie, die Langgriffligkeit auf Homozygotie beruht. Das Beibehalten des 1:1 -Verhältnisses wird durch permanente Rückkreuzungen aufrechterhalten.
Dieses Experiment weist darauf hin, daß es in der Natur keineswegs nur reine Linien gibt und daß viele Phänotypen auf Heterozygotie zurückgeführt werden können. Die dominanten Homozygoten treten unter natürlichen Bedingungen in vielen Arten fast nie in Erscheinung.
Der Unterschied zwischen den beiden Blütentypen beruht nicht nur auf Lang- und Kurzgriffligkeit, vielmehr sind auch die Griffeloberfläche und die Form und Größe des Pollens unterschiedlich. Demnach haben wir es hier zugleich mit einem Beispiel für Pleiotropie zu tun, oder aber mit einer Vielzahl gekoppelter Merkmale (oder beidem).
Wegen der hohen Variabilität der Blütenfarben ist das Löwenmäulchen seit langem eine beliebte Gartenpflanze, und es lag natürlich stets im gärtnerischen Interesse, besonders attraktiv aussehende Formen als reine Linien zu gewinnen. Erste Ansätze hierzu gehen ins 17. Jahrhundert zurück, TOURNEFORT befaßte sich mit ihm um 1700, im 19. Jahrhundert waren zahlreiche Varietäten weit verbreitet; durch Einfuhr und Nutzung neuer Arten wurde zudem die Mannigfaltigkeit der Formen gesteigert. Mehr dazu unter
E. BAUR (1875-1933), erster Direktor des 1927 gegründeten Kaiser-Wilhelm-Instituts für Züchtungsforschung in Müncheberg / Mark (heute: Max-Planck-Institut f. Züchtungsforschung, Köln-Vogelsang) wählte es als eines seiner Hauptversuchsobjekte zur Analyse der Variabilität von Blütenfarben. Hier zunächst aber ein anderes seiner Experimente:
Es gibt eine Form ("aurea") mit goldgelben, statt normal grünen Laubblättern. Kreuzt man solche Pflanzen untereinander, erhält man grüne und aurea-Phänotypen im Verhältnis 1:2. In den Aussaatschalen findet man zusätzlich elfenbeinfarbige Keimlinge, die kurz nach der Keimung eingehen und daher das o.g. Spaltungsverhältnis nicht beeinflussen. E. BAUR schloß daraus, daß die "aurea"-Form heterozygot ist, die grüne dominant homozygot und die elfenbeinfarbigen Keimlinge die rezessiv homozygoten Formen darstellen. Letztere sind nicht lebensfähig und treten daher in ausgewachsenem Zustand nicht in Erscheinung. Die Allelkombination gg ist folglich letal. In diesem Fall ist die Letalität (= Sterblichkeit) physiologisch leicht erklärbar. Das blasse Aussehen beruht offensichtlich auf Chlorophyllabwesenheit, und sobald der Nährstoffvorrat der Samen aufgebraucht ist, wird die Entwicklung abgebrochen, weil sich die Pflanzen nicht weiter ernähren können. Ganz allgemein kann Letalität auf ungünstige Gen- oder Allelkombinationen (Letalitätsfaktoren) zurückgeführt werden und in unterschiedlichen Entwicklungsstadien zum Zuge kommen.
Der amerikanische Genetiker A. J. MANGELSDORF hatte bis 1926 19 voneinander verschiedene Letalitäts- oder Letalfaktoren beim Mais (Zea mays) identifiziert. Umgekehrt ist ein Sortiment solcher Formen ein geeignetes Ausgangsmaterial für entwicklungsphysiologische Studien.
Zeitpunkt der Wirkung einer Reihe von Letalfaktoren (gelbe Querstriche, die Bezeichnungen wären de1, de2, de3......developmental mutant) beim Mais (Nach A. J. MANGELSDORF, 1926)
Eine genaue Analyse (mit modernen Techniken) läßt Schlüsse über Zeitpunkt und Art der einzelnen Genwirkungen zu. Letalität ist naturgemäß ein relativer Begriff. Er besagt lediglich, daß ein Individuum unter gegebenen Umweltsbedingungen nicht lebenstüchtig ist, sich aber unter veränderten Bedingungen ggf. ganz normal entwickeln kann. Chloroplastenfreie Pflanzen beispielsweise können auf zuckerhaltigem Nährsubstrat unter Zusatz von Wuchsstoffen durchaus kultiviert und am Leben erhalten werden.
Wir haben bereits Beispiele für Pleiotropie kennengelernt, ein Gen beeinflußt demnach mehrere Merkmale. Es gibt aber auch den entgegengesetzten Fall, daß mehrere Gene ein Merkmal beeinflussen.
Der schwedische Genetiker H. NILSSON-EHLE (Landwirtschaftliche Forschungsstation Svalöf) berichtete 1909 über eine Kreuzung zwischen einer schwarzkörnigen (genauer gesagt dunkelkörnigen) und einer weißkörnigen (hellen) Hafersorte. In der F1 fand er nur "schwarze" Körner, in der F2 entfielen auf 630 "schwarze" 40 "weiße", was einem Verhältnis von 15:1 entspricht. Er ging nun davon aus, daß zwei voneinander unabhängige Gene (N, M) sowohl jedes für sich als auch beide zusammen die dunkle Kornfarbe bewirken. In der F2 entstehen dadurch nach dem Schema eines dihybriden Erbgangs 15 Individuen, bei denen zumindest eines der Gene in heterozygot dominantem Zustand vorliegt. Alle Genotypen, die mindestens ein dominantes N oder ein dominantes M enthalten, führen zu "Schwarzkörnigkeit"; nur die doppelt rezessiv Homozygoten führen zu "Weißkörnigkeit". Beide Gene wirken gleichsinnig und gleichartig.
Ein vergleichbares Ergebnis erhielt der amerikanische Genetiker G. H. SHULL (Princeton Unversity) bei Kreuzungen zwischen zwei Hirtentäschelkrautarten: Capsella bursa-pastoris und Capsella heegeri. Erstere ist durch herzförmige Schötchen, letztere durch ovale gekennzeichnet. In der Fl sind alle herzförmig, in der F2 kommt ein ovales auf 15 herzförmige. Beide Gene beeinflussen die Fruchtform in gleicher Weise, eines kann das andere gleichwertig vertreten.
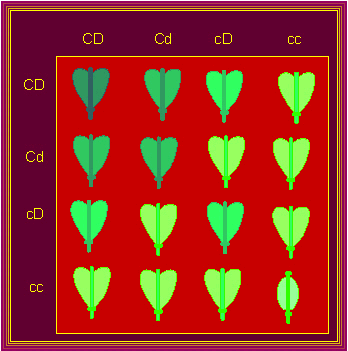
Die Vererbung der Fruchtformen bei Capsella "Punnett-Square". Auf der Abszisse ist der Genotyp der haploiden Gameten des einen, auf der Ordinate der des anderen Elters dargestellt : CD , Cd, cD, cc . - Daraus ergeben sich die Kombinationen (in der ersten Reihe des Quadrats): CDCD, CDCd, CD,cD, Cdcd. Die Kombinationen in der zweiten Reihe wären dann CdCD, CdCd, CdcD, Cdcc, usw. Die Farben im Diagramm symbolisieren die unterschiedlichen Phänotypen. (G. H. SHULL, 1914)
12 + 3 + 1 macht zusammen 16. Schwarzgefärbte Samen der Buschbohne ergeben nach Kreuzung mit bestimmten Sorten weißer Bohnen eine schwarzsamige Fl-Generation, die in der F2 aber nicht nur aus schwarzen und weißen Phänotypen besteht, sondern auch braunsamige Typen enthält. Hier ist das Spaltungsverhältnis 12 schwarz : 3 braun : 1 weiß. Dies ist aber offensichtlich wieder nur eine Variante eines in zwei Merkmalspaaren heterozygoten Bastards, mit dem Unterschied jedoch, daß die beiden voneinander unabhängigen Gene nicht gleichartig sind. Eine biochemische Charakterisierung der Farbstoffzusammensetzung ergab, daß die Dunkelfärbung durch zwei unterschiedliche, dunkle Substanzen hervorgerufen wird. Das dominante Allel des Gens A bewirkt die Bildung eines schwarzen Farbstoffs; das entsprechende Allel von B den braunen. A ist 12mal vertreten. Das trifft zwar auch für B zu, doch wird die Ausprägung des braunen Farbstoffs durch gleichzeitige Anwesenheit des schwarzen (in 9 der 12 Fälle) überdeckt.
Generell spricht man von Epistasie, wenn ein Gen das Sichtbarwerden der Wirkung eines anderen überdeckt. Dieser Begriff gilt meist nur unter bestimmten Voraussetzungen. Analysiert man die Farbstoffzusammensetzung in den einzelnen Samen der Nachkommenschaft mit Hilfe biochemischer (chromatographischer) Methoden, bei denen beide Farbstoffe voneinander getrennt und anschließend identifiziert werden können, erhält man für jeden eine unabhängige Verteilung (Segregation) nach dem MENDELschen Spaltungsverhältnis 3:1 .
Doch liegen die Dinge nicht immer so einfach wie im geschilderten Fall. Epistasie kann sich nämlich auch darin äußern, daß ein weiteres Gen (hypostatisches Gen) überhaupt nicht zur Wirkung gelangt.
Die vorangegangenen Beispiele (15:1, 12:3:1) waren durch Segregation voneinander unabhängiger Gene erklärbar. Ein Verhältnis von 9:7 in der F2 erhält man bei dihybridem Erbgang, bei dem zwei unselbständige Gene komplementär zusammenwirken, d.h., wo jedes ohne das andere nicht in Erscheinung treten kann. So fand KAPPERT, daß bei gewissen Erbsensorten mit dem Merkmal violetthülsig, in entsprechenden Kreuzungen mit grünhülsigen Früchten, in der F2 neun violetthülsige auf sieben grünhülsige Früchte entfielen.
Es genügt also nicht, wenn nur eines der Gene in dominantem Zustand vorliegt. Durch spätere biochemische Analyse solcher Phänotypen konnte eine sinnvolle Erklärung gegeben werden. Die Bildung des violetten Farbstoffs erfolgt nämlich in zwei Schritten. Im ersten Schritt (bedingt durch das Gen V) entsteht aus einer Vorstufe des Farbstoffs ein ebenfalls noch farbloses Zwischenprodukt. Das Gen U bewirkt, daß dieses anschließend in den violetten Farbstoff umgewandelt wird. Wenn V in doppelt rezessivem Zustand vorliegt, kommt U nicht zur Wirkung, weil das Zwischenprodukt fehlt. Wenn U hingegen doppelt rezessiv ist, wird zwar das Zwischenprodukt gebildet, aber die Umwandlung in den Farbstoff unterbleibt.