
Botanik online 1996-2004. Die Seiten werden nicht mehr bearbeitet, sie bleiben als historisches Dokument der botanischen Wissenschaft online erhalten!
Ausschnitt aus dem Leitbündelzylinder einer Kaktee (Opuntia echiops). Bedingt durch die gegenüber den übrigen Geweben verringerte Verwesungsrate verholzter Teile findet man an natürlichen Kakteenstandorten sehr oft auf diese Weise freigelegte verholzte Sproßteile.
Die bisher dargelegten Ausführungen über den Bau von Leitbündeln beruhen auf der Auswertung von Querschnittspräparaten von Stengeln und Wurzeln. Wie wir gesehen haben, eignen sich diese zur Darstellung der Position und der Organisation der Leitbündel (Lage von Xylem, Phloem, Kambium usw.), sie sagen jedoch nichts über die räumliche (dreidimensionale) Anordnung und die Entstehung des Leitbündelsystems aus. Es ist nicht ganz einfach, sich ein anschauliches Bild hierüber zu machen. Serienschnitte und die daraus abgeleitete Rekonstruktion des Leitbündelverlaufs sind zwar ein notwendiges, doch sehr zeitraubendes Verfahren.
Eine weitere Möglichkeit besteht in der Mazeration des Stengels, also praktisch einem kontrollierten Fäulnisprozeß, bei dem man davon ausgeht, daß die Leitbündel die mit am widerstandsfähigsten Gewebe sind und den Mazerationsprozeß weitgehend unbeschadet überstehen.
Trotz methodischer Widrigkeiten sind die genannten Wege mit Erfolg beschritten worden. Von zahlreichen Pflanzenarten aus den verschiedensten Klassen kennen wir heute den Verlauf der Leitbündel; zusätzliche Informationen erhielt man durch die Auswertung von Fossilien.
Einen ersten umfassenden, reich bebilderten Überblick über den Gefäßbündelverlauf findet man in der 1877 erschienenen "Vergleichenden Anatomie der Vegetationsorgane der Phanerogamen und der Farne" des in Straßburg wirkenden Pflanzenanatomen A. de BARY. Er erkannte, daß die Organisation der Leitbündel in den einzelnen Pflanzengruppen auf bestimmte Grundmuster zurückzuführen ist. Von ihm stammen auch die heute noch geläufigen Begriffe axialer Strang (Axialstrang), Blattspur(-strang) und Blattlücke. Heutzutage ist es üblich, von der Stelärtheorie zu sprechen, wenn man sich mit dem Verlauf und der Entstehung von Leitbündeln auseinandersetzt.
Das Konzept der Stele geht auf P. van TIEGHEM, Professor am Musée d'Histoire Naturelle de Paris und seinen Schüler H. DOULIOT (1886) zurück. Ihre Vorschläge wurden vielfach überarbeitet und abgewandelt. Die von van TIEGHEM vorgeschlagene Terminologie ist heute nur noch von historischem Interesse.
Besonders um die Jahrhundertwende gab es kaum einen namhaften Morphologen, der sich nicht mit dieser Problematik befaßte. Nennenswert sind vor allem E. STRASBURGER (Professor der Botanik an der Universität Bonn) mit seiner 1891 erschienenen Arbeit "Über den Bau und die Vorrichtungen der Leitungsbahnen in der Pflanze", E. C. JEFFREY (University of Toronto, später Harvard University), der 1898 eine Reihe neuer Begriffe definierte, u.a. Protostele und Siphonostele (mit denen wir uns noch ausgiebig befassen werden), sowie die Engländer G. BREBNER (1902) und F. O. BOWER.
Der unterschiedliche Gebrauch von Begriffen und eine eigenständige Nomenklatur trugen gerade in jener Zeit mehr zur Verwirrung als zur Klärung bei. Um 1910 stabilisierte sich die Lage, und man einigte sich auf Definitionen, die als "British System" in die Literatur eingegangen sind.
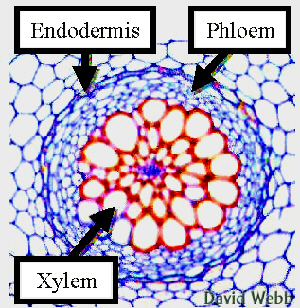
Unter Stelen faßt man das Leitbündelsystem, einschließlich des mit ihm assoziierten Gewebes sowie das vom Leitbündelzylinder umgebene Mark zusammen. Der Ausdruck Stele entstammt, wie so viele Ausdrücke der Pflanzenanatomie dem Griechischen und bedeutet Säule. Die Stelärtheorie besagt, daß der primäre Pflanzenkörper im Stengel und in der Wurzel nach dem gleichen Prinzip konstruiert ist, weil beide eine zentrale Säule (Stele) enthalten, die in primäres Rindengewebe eingebettet ist. Die Stelärtheorie befaßt sich nicht mit dem sekundären Dickenwachstum. Das steläre Muster kann als ein konservatives Merkmal angesehen werden, dessen Komplexität im Verlauf der Evolution zunahm.
Die Protostele, ein einfach organisiertes Leitsystem. Die Protostele ist ein einfacher, unverzweigter, zentral gelegener Axialstrang aus Xylem, das vom Phloem ummantelt ist oder bei dem Phloemanteile in das Xylem eingestreut sind. Eine Protostele enthält kein Mark. Man findet sie bei den primitiven Gefäßpflanzen, z.B. der fossilen Rhynia, sowie in jungen Farnsprossen, in Stengeln einiger einfacher Wasserpflanzen und in den meisten Wurzeln der Angiospermen. Eine Variante, die Plectostele ist für die Lycopodien (Bärlappgewächse) typisch
 Einfache Stele aus der Wurzel von Lycopodium. |
 |
Die Siphonostele im weitesten Sinne. Die Siphonostele stellt den Prototyp der Leitbündelsysteme der Farne und aller weiter entwickelten Gefäßpflanzen (Samenpflanzen) dar. Sie besteht aus mehreren axialen Leitbündeln, die im Stengel in Form eines Hohlzylinders angeordnet sind und ein Markgewebe umschließen.
 Aktinostele von Psilotum: Das sternförmig angeordnete Xylem (Actino = Star) ist von Phloem umgeben. |
 |
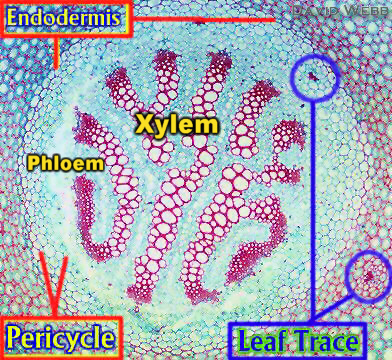
Von den axialen Leitbündeln zweigen Blatt- und Zweigspuren ab, die sich in die Blätter, bzw. Seitenzweige hinein erstrecken. Eine Blattspur beginnt stets am Abzweigungspunkt und reicht bis zur Blattbasis (Blattansatzstelle). Ein Blatt kann durch eine oder mehrere Blattspuren versorgt werden. Die Axialbündel (axiale Leitbündel) erstrecken sich über die gesamte Länge eines Stengelabschnitts hinweg. Vergegenwärtigt man sich, wie Leitbündel im Verlauf der Ontogenese entstehen, wird sofort klar, daß sie aus Segmenten zusammengesetzt sind, die ursprünglich als Blattspuren angelegt waren.
Der Übergang Blattspur zu Leitbündel geschieht, sobald eine apikalwärts angelegte Blattspur den Kontakt zum bestehenden Leitbündelsystem aufnimmt.
Axialbündel sind in der Regel durch Querverbindungen untereinander vernetzt, wodurch ein kommunizierendes Leitungssystem zustande kommt (K. J. DORMER, 1954). Die Bereiche, in denen Blattspuren abzweigen, sind die Knoten oder Nodien und die zwischen ihnen liegenden Abschnitte die Internodien. Die Struktur der Knoten in den Stengeln der Dikotyledonen ist relativ einfach, komplexe Vernetzungen treten vorwiegend bei den Monokotyledonen auf.
Einige Beispiele: Einfache Verhältnisse kommen bei primitiven fossilen Vorläufern der Gymnospermen (Progymnospermen) vor, so z.B. bei der Art Callixylon brownii, deren Leitbündelsystem C. B. BECK 1979 rekonstruiert hat. Es zeigt im wesentlichen parallel geordnete Leitbündel, zwischen denen es nur wenige Verknüpfungen (Anastomosen) gibt. Eine beträchtliche Steigerung der Komplexität des Verzweigungsmusters tritt bei den Gymnospermen auf.
Man unterscheidet zwischen den offenen Leitbündelsystemen und den geschlossenen, bei denen die Blattspuren durch Anastomosen untereinander in Verbindung stehen. Bei Ginkgo biloba wird jedes Blatt durch zwei Blattspuren versorgt, die sich von getrennten, aber benachbarten Axialbündeln ableiten. Bei den Cycadales (z.B. Dioon spinulosum) sind fünf Blattspuren pro Blatt vorhanden, die entfernt liegenden Axialsträngen entspringen. Die Blattspuren liegen dem Leitbündelzylinder zunächst ringförmig an, bevor sie vereint in Richtung Blatt abzweigen (H. A. DORETY, 1919). Hier haben wir es bereits mit einer recht komplexen Organisation der Knotenstruktur zu tun.

Angiospermen zeichnen sich durch eine hohe Variabilität in der Konstruktion der Leitbündelzylinder aus, obwohl die Grundmuster weitgehend identisch sind. Es gibt jedoch einen gravierenden Unterschied zwischen den Dikotyledonen und den Monokotyledonen. Die Situation bei den Dikotyledonen steht dem Grundmuster am nächsten, ist am übersichtlichsten und wird daher zuerst besprochen:
Die Variationen beruhen auf Richtung und Ganghöhe der Schraube, Anzahl der Spuren pro Blatt und Natur der Blattansatzstelle. In diesem Zusammenhang muß der Begriff Blattlücke fallen, der sich auf eine Aussparung in Leitbündelzylindern oberhalb der Blattansatzstelle bezieht.
 Stelentypen.
Links: Siphonostele ohne Blattlücken. Mitte und rechts:
Siphonostelen mit Blattspuren (Nach Y. OGURA, 1938).
Stelentypen.
Links: Siphonostele ohne Blattlücken. Mitte und rechts:
Siphonostelen mit Blattspuren (Nach Y. OGURA, 1938).
In der Regel enthalten Angiospermen fünf Leitbündelstränge. Man betrachtet diese Zahl als Grundeinheit, von der Varianten mit mehr oder weniger Strängen abgeleitet sind. In 91 Prozent aller Arten kommen offene Systeme vor, geschlossene (verschiedene Blattspuren und Blattspurkomplexe miteinander verbunden) bei den übrigen (sowohl bei Kräutern als auch bei Holzpflanzen). Die Zahl der Leitbündel pro Leitbündelstrang ist dort geradzahlig mit vier als Grundzahl.
Vielfach sind Blattlücken sehr groß (z.B. bei vielen Angiospermen, unter anderem bei den Kakteen, aber auch bei Cycadeen und Farnen). Der Leitbündelzylinder sieht daher netzartig strukturiert aus.
Zusammenfassend können im Verlauf der Evolution der Samenpflanzen folgende Trends ausgemacht werden (C. B. BECK et al., 1982):
Offene Leitbündelsysteme sind primitiv, geschlossene abgeleitet.
Der unilakunare Typ (nur eine Blattlücke) ist primitiv, multilakunare gelten als abgeleitet. Ausnahmen, Übergänge und Rückentwicklungen findet man bei vielen Dikotyledonen. Oft findet man zwei oder mehrere Typen innerhalb einer Pflanze. Das unilakunare Knotensystem erscheint in älteren geologischen Schichten als das trilakunare. Es ist der häufigste Typ bei primitiven Samenpflanzen.
Leitbündelsysteme mit fünf Strängen sind vermutlich primitiv, diejenigen mit mehr oder weniger Strängen abgeleitet.
Stelen (Leitbündelsäulen) mit Blattspuren, die in Längsrichtung (longitudinal) ein oder nur wenige Internodien durchlaufen, können als primitiv, solche, die viele Internodien durchlaufen, als abgeleitet angesehen werden.
Bei Samenpflanzen gilt eine helicale (schraubige) Anordnung der Blattanlagen als primitiver als andersartige Anordnungen.
Bei den Monokotyledonen scheinen die Leitbündel über den gesamten Stengel verteilt zu sein, wobei sie nahe der Peripherie einander genähert sind. Betrachtet man den Verlauf eines einzelnen Axialbündels, erkennt man eine klare schraubige Form. Niemals liegt ein Leitbündel nur zentral oder nur peripher. Je nachdem, wo man den Stengel anschneidet, trifft man es daher mal im Zentrum, mal in peripherer Lage an. In regelmäßigen Abständen zweigen Blattspuren ab. Auch sie verlaufen über lange Strecken hinweg im Stengel, liegen in der Regel jedoch peripher. Dies ist auch der Hauptgrund, weshalb man in Querschnitten gerade dort besonders viele Leitbündel sieht. Ein Teil von ihnen stellt Axialstränge, ein anderer Blattspuren dar. In Querschnitten sind sie meist nicht voneinander unterscheidbar, obwohl Axialstränge in der Regel einen etwas größeren Durchmesser haben. Axialbündel können durch Brückenbündel untereinander verknüpft sein.
Unter Berücksichtigung dieses Bauplans erscheint die durch Betrachtung von Querschnitten gewonnene Aussage, die Leitbündel seien im Stengel verstreut angeordnet, irreführend. Der Konstruktionsplan veranschaulicht, daß hier, wie auch bei den Dikotyledonen (und den Gymnospermen), eine klare Rotationsymmetrie vorliegt, bei der die Form des Leitbündelzylinders jedoch weitgehend abgewandelt ist, im Querschnitt sogar aufgehoben erscheint. Jedes Leitbündel wird während der Ontogenese in peripherer Position angelegt, und erst während des nachfolgenden Wachstums geraten einzelne Abschnitte streckenweise ins Zentrum des Stengels.