Botanik online 1996-2004. Die Seiten werden nicht mehr bearbeitet, sie bleiben als historisches Dokument der botanischen Wissenschaft online erhalten!
Dazu J. v. SACHS:
"Die Gesamtheit aller Gewebemassen, welche von dem Hautgewebe (Anm.: der Epidermis) umschlossen und von den Gefäßbündeln durchzogen sind, fasse ich unter dem Ausdruck Grundgewebe zusammen, wie ich es in der 1. Auflage meines Lehrbuchs 1868 zuerst charakterisiert habe. Bei jüngeren, noch saftigen, nur von der Epidermis bedeckten Organen, deren Gefäßbündel noch nicht durch nachträgliches Dickenwachstum deformiert worden sind, bei Organen überhaupt, in welchen die Bildung echten Holzes und sekundärer Rinde noch nicht eingetreten ist, besteht die Hauptmasse aus Grundgewebe. Zu klarster Anschauung gelangt dasselbe vielleicht bei Betrachtung eines Apfels, dessen ganze genießbare Substanz daraus besteht.. ... Die verbreitetste und, wie man wohl annehmen darf, ursprünglichste Form des Grundgewebes ist das gewöhnliche, dünnwandige Parenchym."
Parenchymatische Zellen sind durchweg lebend, meist isodiametrisch, seltener gestreckt. Das Mark der Sprosse, die Speichergewebe der Früchte, Samen, der Wurzel und anderer unterirdischer Organe sind ebenso als Parenchyme zu betrachten wie das Mesophyll (Assimilationsgewebe der Laubblätter). Wegen der fundamentalen Bedeutung für die Assimilation und damit für die gesamte Ernährung der Pflanze ist es zweckmäßig, das Mesophyll in einem gesonderten Abschnitt (im Anschluß an die Besprechung des weniger spezialisierten Parenchyms) zu behandeln.
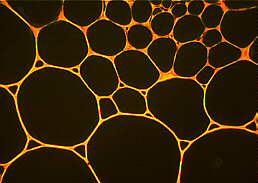
Parenchymatische Zellen und relativ große Interzellularen in einem Stengelquerschnitt von Geum urbanum . Fluorochromierung mit Coriphosphin. Die Mittellamelle erscheint hier als rote Fluoreszenz.
Oft kommt den Parenchymen anscheinend nur eine einfache Füllfunktion zu. Wie schon der Name Grundgewebe ausdrückt, stellt es die Masse der Zellen, in welches spezialisierte Gewebe, wie Leitgewebe und oft auch die Samen eingebettet sind.
Parenchymzellen sind weder morphologisch noch physiologisch spezialisiert. Vielfach enthalten sie Chloroplasten, manchmal, vor allem dort, wo sie sich unter Lichtausschluß entwickelt haben, Leukoplasten oder unvollständig strukturierte Chloroplasten. Obwohl in ausdifferenzierten Parenchymen keine oder nur vereinzelte Zellteilungen stattfinden, behalten die Zellen ihre Teilungsbereitschaft, was allein schon darin zum Ausdruck kommt, daß sie die Hauptmenge der an der Wundheilung und Regeneration von Pflanzenteilen beteiligten Zellen stellen. Man könnte demnach vielleicht sagen, daß sich die Zellen in Wartestellung befinden. Sie bilden einen Zellpool (einen Zellvorrat), der bei Bedarf, d.h., während der normalen Ontogenese und ebenso bei außergewöhnlichen äußeren Anlässen aktiviert werden kann.
Das Meristem haben wir als das Teilungsgewebe der Pftanzen charakterisiert. Das Parenchym könnte man entsprechend als ein Ausgangsgewebe beschreiben, von dem aus sich während der Ontogenese die unterschiedlichsten Zelltypen ableiten. Auf die Bedeutung der Position einer Zelle in Beziehung zu den übrigen wird im Thema Zellen und Gewebe hingewiesen, von ihr hängt es ab, ob sich eine Zelle teilt oder nicht (Positionsinformation). Auch bei den parenchymatischen Zellen entscheidet die Lage einer Zelle über ihr weiteres Schicksal und darüber, in welche Richtung sich die Zelle (oder eine Gruppe benachbarter Zellen) weiterentwickelt.
Parenchymatische Gewebe (z.B. aus dem Mark des Sprosses) können auf geeigneten synthetischen Nährmedien (Nährböden) kultiviert und dadurch die Zellen zu Teilungen angeregt werden. Die Teilungskapazität wird über Jahrzehnte beibehalten, wenn Teile der Kulturen in regelmäßigen Zeitabständen auf ein frisches Medium überführt werden. Die Zellen verbleiben in einem weitgehend unspezialisierten Zustand. Sie bilden Aggregate aus, die als Kalli (sing. Kallus) bezeichnet werden; die Kulturen selbst nennt man Gewebe- oder Kalluskulturen. Hält man sie bei Dunkelheit, bleiben die Kalli farblos; bei Belichtung ergrünen sie meist. Durch Zusatz geeigneter Wuchsstoffe (Phytohormone) können Sproß- und/oder Wurzelbildung induziert werden.
Im Stoffwechsel der Pflanzen entsteht eine Vielzahl spezifischer Substanzen, von denen ein Teil gespeichert und zu einem späteren Zeitpunkt gegebenenfalls genutzt wird. Es sind wiederum vorwiegend parenchymatische Zellen in den verschiedensten ober- und unterirdischen Organen, die als Depot dienen. Mit der Chemie der gespeicherten Substanzen werden wir uns später noch ausgiebig befassen. Hier sei nur vermerkt, daß grundsätzlich zwischen kleinen und großen Molekülen (Makromolekülen) zu unterscheiden ist. Zu den kleinen rechnet man - außer den anorganischen Ionen - organische Säuren (bzw. ihre Salze), Zucker, eine Anzahl stickstoffhaltiger Verbindungen (z.B. Aminosäuren, Alkaloide) u.a., zu den Makromolekülen die Stärke sowie die Proteine. Kleine Moleküle werden meist in gelöster Form in den Vakuolen gespeichert. Gelegentlich kristallisieren sie aus, und die Kristalle werden je nach Art entweder in der Vakuole oder im Plasma abgelagert. Makromoleküle gelangen nur selten in die zentrale Vakuole. Sie werden als mikroskopisch sichtbare Aggregate (Proteinkörper, Stärkekörner) im Plasma deponiert.
Zucker und Stärke gehören zu den wichtigsten Primärprodukten der Photosynthese. Bekanntlich sind verschiedenste Zucker in mitunter beträchtlichen Mengen im Parenchym von Früchten (dem Fruchtfleisch) enthalten. Die gleichzeitige Speicherung organischer Säuren (in den gleichen Zellen) verleiht ihnen den artspezifischen, oft süß-sauren Geschmack. Stärkedepots findet man in Samen (im Endosperm, ebenfalls einem parenchymatischen Gewebe), in Wurzeln, Wurzelknollen (Dahlien, Batate), Sproßknollen (Kartoffel) u.a.
Proteine werden ebenfalls oft in Samen gespeichert, zumeist in darauf spezialisierten Zellen. In einem Weizenkorn findet man beispielsweise in den zentral gelegenen Endospermzellen Stärke und in weiter außen liegenden Protein in Form sogenannter Aleuronkörner. Bei den Proteinen handelt es sich um Speicherproteine, die einer bestimmten Proteinklasse angehören. Ihre chemische Zusammensetzung und Struktur ist artspezifisch.
Beim Auskeimen der Samen werden die im Endosperm gelagerten Makromoleküle (Reservestoffe) partiell degradiert und dienen dem Aufbau des Keimlings. Nach Mobilisierung ihrer Inhaltsstoffe stirbt dieses Gewebe in der Regel ab.
Neben den eben genannten Verbindungen speichern Parenchymzellen große Mengen an Wasser. Besonders augenscheinlich in Geweben sukkulenter (dickfleischiger) Pflanzen: Kakteen, Aloe, Agave, Aizoaceae ( = Mittagsblumengewächse ). In den einzelnen Abschnitten eines Bambushalmes wurde eine direkte (lineare) Korrelation zwischen prozentualem Anteil an Parenchymgewebe und Wassergehalt ermittelt.
Aber auch bestimmte Teile der Pflanzen unserer Breiten sind wasserhaltiger als die übrigen: dazu zählen z.B. das Fruchtfleisch, die Zwiebeln, die Knospen, alle "fleischigen" Verdickungen oberirdischer Teile usw.
Die Kapazität, in den Vakuolen Wasser zu speichern, ist von der
Molarität der darin gelösten Substanzen abhängig. Man spricht
dabei vom osmotischen Wert (Osmolarität).
Im vorangegangenen Abschnitt haben wir die Vakuolen parenchymatischer Zellen als Depots für eine Reihe kleiner Moleküle kennengelernt. Aus der eben postulierten physikochemischen Bedingung folgt zwingend, daß gerade solche Zellen für eine hohe Wasseraufnahmerate prädestiniert sind. Man erkennt das allein schon daran, daß reife Früchte stets prall sind; in überreifen kann der hohe osmotische Druck sogar zu ihrem Platzen führen.
In den meisten parenchymatischen Geweben, vor allem denen mit Speicherfunktion, gibt es nur kleine, teilweise sogar gar keine Interzellularen. Das ist zum einen durch den hohen osmotischen Druck (=Turgor) der einzelnen Zellen, zum anderen durch deren dünne Zellwände erklärbar, die der Ausdehnung des Zellinhalts nur wenig Widerstand entgegensetzen und es dadurch gar nicht erst zur Bildung von Interzellularen kommen lassen.
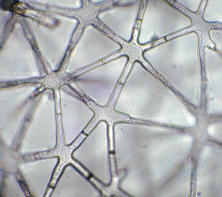
Anders als gerade beschrieben sieht es bei einem speziellen Typ parenchymatischer Zellen aus, der sich durch stärkere Wände auszeichnet. Durch Auseinanderweichen solcher Zellen oder durch Reißen der Gewebe entstehen lufterfüllte Hohlräume. Die durch Reißen entstehenden Hohlräume sind im Sproß und in den Wurzeln meist tangential oder radial angelegt. Die Ursache für das Reißen ist in Spannungsdifferenzen zwischen benachbarten, sich unterschiedlich vergrößernden Geweben zu suchen. Beim Auseinanderweichen, beruhend auf asymmetrischem Streckungswachstum, entstehen unregelmäßig geformte Zellen, zwischen denen sich umfangreiche Interzellularsysteme ausbilden. Beispielhaft dafür ist das Schwammparenchym der Laubblätter. Ausgedehnte Durchlüftungsgewebe findet man aber auch im Mark vieler Monokotyledonen feuchter Standorte, z.B. bei den Binsen. Wegen der dort auftretenden spezifischen Zellform wird der Gewebetyp als Sternparenchym bezeichnet.
Sternparenchym aus dem Mark des Stengels von Binsen (Juncus). Extrem große Interzellularen. Das Parenchym dient hier als ein Durchlüftungsgewebe
(Aufn.: W. KASPRIK).
In submers (untergetaucht) wachsenden Sprossen vieler Angiospermen, so z.B. bei der Weißen und der Gelben Teichrose kommen im Parenchym große, schon mit bloßem Auge erkennbare Luftkanäle vor, die durch einfache Zellschichten voneinander getrennt sind. Diese wabenartige Konstruktion bedingt zweierlei: Zum einen hohe Festigkeit bei nur geringem Materialeinsatz. Dieses Prinzip wird übrigens auch im technologischen Bereich erfolgreich angewandt. Zum anderen wird durch das geringe spezifische Gewicht ein Auftrieb erzeugt, der der Schwimmfähigkeit der Sprosse zugute kommt.
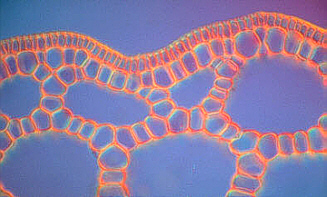
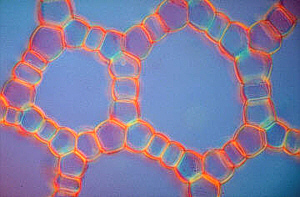
Aerenchym in Stengelquerschnitten von
Hippuris vulgaris