C. References
XIII. Botanical fallouts and Biological Significance of the Chl Biosynthetic Heterogeneity
A. Botanical fallout of the Chl a Biosynthetic Heterogeneity: Classification of Green Plants into Various Greening Groups
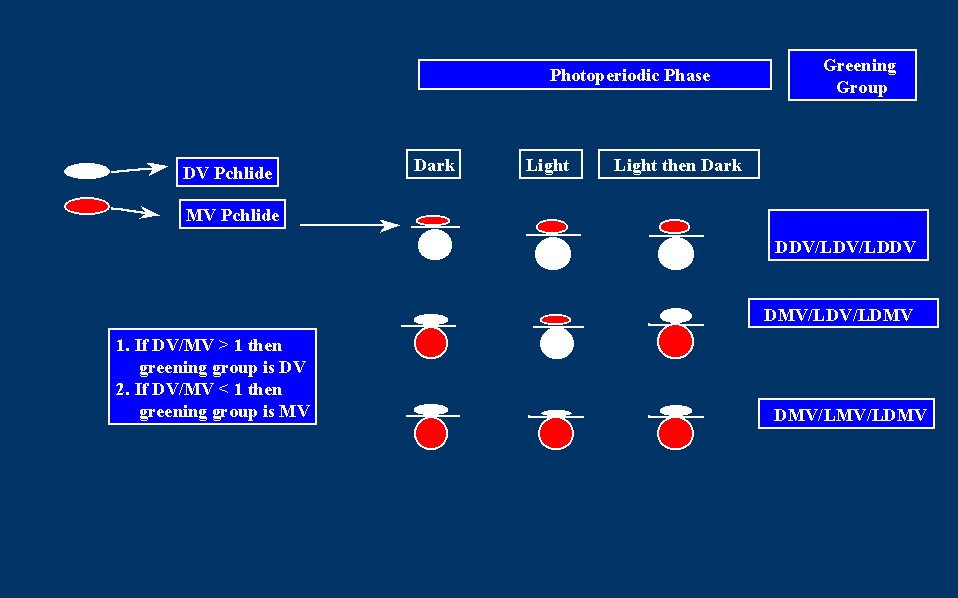
Fig. 6. Greening Group Affiliation of Green Plants
If the DV/MV Pchlide a ratio for a plant species is larger than 1, for a particular phase of the photoperiod, then the plant is considered to be DV for that phase of the photoperiod. If the ratio is less than one, the plant is considered to be MV for that phase of the photoperiod. Plants that are DV during the dark phase of the photoperiod, are referred to as DDV. Plants that are MV during the dark phase of the photoperiod, are referred to as DMV. Likewise Plants that are DV during the light phase of the photoperiod, are referred to as LDV. Plants that are MV during the light phase of the photoperiod, are referred to as LMV. Plants that accumulate DV Pchlide a immediately after returning them from light to darkness are referred to as LDDV. Plants that accumulate MV Pchlide a immediately after returning them from light to darkness are referred to as LDMV. For the whole photoperiod, plants can be DDV/LDV/LDDV, DMV/LDV/LDMV or DMV/LMV/LDMV. The DDV/LMV group has not been observed under natural conditions.
Biosynthetic heterogeneity refers to the biosynthesis of a particular metabolite by an organelle, tissue or organism via multiple biosynthetic routes (see Heme and Chl a Biosynthetic Pathways). It has been well documented in delta-aminolevulinic acid (ALA), chlorophyll (Chl) a and vitamin B12 biosynthesis (Arigoni, 1994; Rebeiz et al, 1994; Scott, 1994). It has been demonstrated that in green plants, Chl a is formed via parallel biosynthetic routes, namely (a) a DV Chl a biosynthetic route which originates in the DV Proto pool, (b) a MV route which originates in the DV Mg-Proto pool, and (c) mixed DV-MV routes which bifurcate at the levels of DV Pchlide a, and DV Chlide a (Fig. 2) (Rebeiz et al, 1994; Leeper, 1991; Richards, 1993; Kim and Rebeiz, 1996). Intermediates of the DV carboxylic route consist of dicarboxylic and monocarboxylic tetrapyrroles with vinyl groups at positions 2 and 4 of the macrocycle, such as DV protoporphyrin IX (Proto), DV Mg-Proto, DV Mg-Proto monomethyl ester (Mpe), DV Pchlide a, and DV Chlide a (Fig. 2). The MV carboxylic routes involve dicarboxylic and monocarboxylic tetrapyrroles including, MV Mg-Proto, MV Mpe, MV Pchlide a, and MV Chlide a, which have one vinyl and one ethyl group at positions 2 and 4 of the macrocycle, respectively (Fig. 2). The mixed DV-MV carboxylic routes involve monocarboxylic tetrapyrroles such as DV and MV Pchlide a and Chlide a.
In higher plants, the end product of the Chl a biosynthetic heterogeneity is invariably MV Chl a and b, with the only known exception of a lethal maize mutant (Bazzaz, 1981) which forms only DV Chl a and b. However, in the prochlorophyte picoplankton of the subtropical waters of the North Atlantic as well as in the picoplankton of the euphotic zone of the world tropical and temperate oceans, and the Mediterranean sea, DV Chl a and b are the predominant Chl species (Veldhuis and Kraay, 1990; Chisholm et al, Goerike and Repeta, 1992; Chisholm et al, 1992). It has been proposed that in green plants, the multiplicity of Chl a biosynthetic routes, produces different pools of MV Chl a, complexed to different pigment-proteins at specific sites of the photosynthetic membranes (Rebeiz et al, 1994; Rebeiz et al; 1983). The DV and MV Chl a biosynthetic routes are linked at the level of DV Mg-Proto (Kim and Rebeiz, 1996), DV Pchlide a (Tripathy and Rebeiz, 1988), and DV Chlide a (Parham and Rebeiz, 1992, 1995) by [4-vinyl] reductase(s) that convert the 4-vinyl group at position 4 to ethyl, thus converting a DV tetrapyrrole to a MV tetrapyrrole. Ellsworth and Hsing (Ellsworth and Hsing, 1974) also reported that DV Mpe could be converted to MV Mpe by an NADH-dependent enzyme. However, so far no one including us has been able to duplicate their results. Whyte and Griffiths (1993) have interpreted the accumulation of DV and MV Pchlide a in term of a dual pathway with a single vinyl reductase of broad specificity. In their scheme the major route converts DV Pchlide a to MV Chl a via DV Chlide a, and MV Chlide a. The minor route consists of the formation of MV Chlide a, and by inference MV Chl a via DV Pchlide a, which is converted to MV Pchlide a by the nonspecific vinyl reductase. This hypothesis is not compatible with the following observations: (a) It has been demonstrated that during DV and MV Pchlide a biosynthesis, only a fraction of the MV Pchlide a pool can be formed by reduction of DV Pchlide a (Tripathy and Rebeiz, 1988), and (b) In Rhodobacter capsulatus in which the bchJ gene which codes for DV Pchlide a reductase (4VpideR) has been deleted, in addition to the accumulation of DV Pchlide a, accumulation of MV Mg-Proto, its monoester (precursors of Pchlide a), and MV Pchlide a have been observed (Suzuki and Bauer, 1995). This in turn indicates that at least one separate [4-vinyl] reductase exists which acts prior to DV Pchlide a and DV Chlide a vinyl reduction. This enzyme would be responsible for the accumulation of MV Mg protoporphyrins in plants (Belanger and Rebeiz, 1982), as well as for MV Pchlide a formation in the absence of 4VpideR. Very recently the Chl a biosynthetic heterogeneity has been extended to the level of Pchlide a photoreduction, by the discovery of a second Pchlide a oxidoreductase (POR), in addition to the conventional POR of etiolated tissues (Armstrong et al, 1995; Holtorf et al, 1995). One, POR-A, is the conventional photoenzyme, which occurs in etiolated tissues but disappears during greening. The second, POR-B is present throughout angiosperm development. Armstrong et al (Armstrong et al, 1995) have suggested that POR-A performs a specialized function restricted to the initial stages of greening, while POR-B is involved in maintaining Chl levels throughout angiosperm development. On the basis of the DV-MV biosynthetic heterogeneity (see above), green plants have been classified into three different greening groups depending upon MV or DV Pchlide a accumulation during the dark and light phases of the photoperiod (Carey and Rebeiz, 1985; Ioannides, et al, 1994; Shioi and Takamiya, 1992). DDV-LDV plants, such as Pogostemon cablin, cucumber (Cucumis sativus), velvetleaf (Abutilon theophrastii), common morningglory (Ipomea purpurea), prickly sida (Sida spinosa), and mustard (Brassica nigra), accumulate mainly DV Pchlide a at night and in daytime. So far, all representative primitive plant species, including algae, bryophytes, ferns, and gymnosperms, fall in this greening group. This has led Ioannides et al (1994) to propose that this greening group is evolutionary ancestral. DMV-LMV) plants, such as barnyardgrass (Echinochloa crus-galli) and johnsongrass (Sorghum halepense), accumulate mainly MV Pchlide a at night and in daytime. This greening group comprises a small number of plants, and evolutionary studies suggest that it is derived (Ioannides et al, 1994). Finally, DMV-LDV plants such as French bean (Phaseolus vulgaris L.), corn (Zea mays), wheat (Triticum aestivum), wild oat (Avena fatua), barley (Hordeum vulgare), soybean (Glycine max), lambsquarter (Chenopodium album), jimsonweed (Datura strmonium), redroot pigweed (Amaranthus retroflexus), cocklebur (Xanthium stumarium), etc., accumulate MV Pchlide a at night. In daytime they accumulate mainly DV Pchlide a. This greening group comprises by far the largest number of plant species so far surveyed, and evolutionary studies suggest that it is evolutionary intermediate (Ioannides et al, 1994). Plant species of major agronomic importance belong to this group (Ioannides et al, 1994). Recently it has been demonstrated, that in addition to differences in the levels of DV or MV Pchlide a accumulation during the dark and light phases of the photoperiod, the three greening groups of plants can be subdivided into subgroups which differ by the dark-rates of DV or MV Pchlide a regeneration when the light phase of the photoperiod is interrupted by a brief period of darkness (Abd El Mageed et al, 1997). Interruption of the light phase of the photoperiod by a brief period of darkness (LD condition) revealed predominance of different Chl a biosynthetic routes, depending upon the greening group affiliation of the plant species. For example, in DMV-LDV and DMV-LMV plants, the predominant Chl a biosynthetic routes under the LD condition appear to be the MV Chl a biosynthetic route and/or a mixed DV-MV Chl a biosynthetic route which bifurcates at the level of DV Pchlide a (Fig. 2). On the basis of DV and MV Pchlide a accumulation rates after re-darkening, this greening group is designated as a light-dark MV (LDMV) subgroup. In DDV-LDV plants, the predominant LD Chl a biosynthetic routes appear to be the DV Chl a biosynthetic route and/or a mixed DV-MV Chl a biosynthetic route which bifurcates at the level of DV Chlide a (Fig. 2). This greening group is designated as a light-dark DV (LDDV) subgroup. It is proposed that upon inhibiting the conversion of Pchlide a to Chl a by interruption of the light phase of the photoperiod by a brief period of darkness, the rates of DV and MV Pchlide a regeneration reflect the carryover rates of DV and MV Pchlide a biosynthesis in the light, instead of reflecting a differential use of DV and MV carboxylic biosynthetic rates in darkness. It was also shown that in LDMV plants, MV Chlide a and MV Chl a are formed without the participation of [4-vinyl] Chlide a reductase. On the basis of recently published evidence, it was also argued that Pchlide oxidoreductase-A (POR-A) may be active in LDDV plants, while POR-B may predominate in LDMV plant species.B. Biological Significance of the Chl Biosynthetic Heterogeneity:
Natural selection has often produced multiple (bio)chemical and physical ways of conveying the same message. Genetic redundency achieves the same purpose. It is therefore logical to thing of the Chl biosynthetic heterogeneity as a hedge against lethal mutations. It is also possible that via natural selection, Chl biosynthetic heterogeneity has imparted an evolutionary advantage to higher plants. As was just described, the preferential operation of the MV or DV Chl a monocarboxylic biosynthetic routes during the formation of Chl a in higher plants is a species-dependent phenomenon with evolutionary significance (Ioannides et al, 1994; Abd El Mageed et al, 1997). Accordingly, evolution under domestication, as manifested by selection for higher plant yields, may favor either one of the two routes. It was shown for example (Fasoula et al, 1996), that in spring wheat (Triticum aestivum L.) and corn (Zea mays L.), plants that use preferentially the more evolved MV Chl a biosynthetic route at night, selection for yield does not favor the DV route, whereas in certain cases the MV route is enhanced.References
- Arigoni, D., (1994) Summing up. In: The Biosynthesis of the Tetrapyrrole Pigments. Ciba Symposium 180 pp 337-338. John Wiley, New York.
- Rebeiz, C. A., R. Parham, D. A. Fasoula, and I. M. Ioannides, (1994). Chlorophyll biosynthetic heterogeneity. In: The Biosynthesis of the Tetrapyrrole Pigments. Ciba Symposium 180 pp 177-193. John Wiley, New York.
- Scott, A. I., (1994). Recent studies of the enzymically controlled steps in B12 Biosynthesis. In: The Biosynthesis of the Tetrapyrrole Pigments. Ciba Symposium 180 pp 285-308. John Wiley & Sons, New York.
- Leeper, F., (1991). Intermediate steps in the biosynthesis of chlorophylls. In: Chlorophylls. (H. Scheer, ed.), pp 407-431. CRC Press, Boca Raton.
- Richards, W., (1993). Biosynthesis of the chlorophyll chromophore of pigmented thylakoid proteins. In: Pigment Protein Complexes in Plastid Synthesis and Assembly. ( C. Sundqvist and M. Ryberg, eds.), pp 91-178. Academic Press, New York.
- Kim, J. S., and C. A. Rebeiz, (1996). Origin of chlorophyll a biosynthetic heterogeneity in higher plants. J. Biochem. Mol. Biol. 29: 327-334.
- Bazzaz, M. B., (1981). New chlorophyll chromophores isolated from a chlorophyll deficient mutant of maize. Photobiochem. Photobiosphys 2:199-207.
- Veldhuis, M. J. W., and G. W. Kraay, (1990). Vertical distribution of pigment composition of a picoplanktonic prochlorophyte in the subtropical north Atlantic: A combined study of pigments and flow cytometry. Mar. Ecol. Progr Ser. 68: 121-127.
- Chisholm, S. W., R. J. Olson, E. R. Zettler, R. Goericke, J. B. Waterbury, and N. A. Welschmeyer, (1990). A novel free-living prochlorophyte abundant in the oceanic euphotic zone. Nature. 334: 340-343.
- Goerike, R., and D. Repeta, (1992). The pigments of Prochlorococcus marinus. The presence of divinyl-chlorophyll a and b in a marine prochlorophyte. Limnol. Oceanogr. 37: 425-433.
- Chisholm, S. W., S. Frankel, R. Goerike, R. Olson, R. Palenic, B. Urbach, J. Waterbury, and E. Zettler, (1992). Prochlorococcus marinus nov.gen. sp.: an oxyphototropic marine prokaryote containing divinyl chlorophyll a and b. Arch. Mikrobiol. 157: 297-300.
- Rebeiz, C. A., S. M. Wu, M. Kuhadje, H. Daniell, and E. J. Perkins, (1983). Chlorophyll a biosynthetic routes and chlorophyll a chemical heterogeneity. Mol. Cell. Biochem. 58: 97-125.
- Tripathy, B. C., and C. A. Rebeiz, (1988). Chloroplast Biogenesis 60. Conversion of divinyl protochlorophyllide to monovinyl protochlorophyllide in green(ing) barley, a dark monovinyl/light divinyl plant species. Plant Physiol. 87: 89-94.
- Parham, R., and C. A. Rebeiz, (1992). Chloroplast Biogenesis: [4-Vinyl] chlorophyllide a reductase is a divinyl chlorophyllide a-specific NADPH-dependent enzyme. Biochemistry. 31: 8460-8464.
- Parham, R., and C. A. Rebeiz, (1995). Chloroplast Biogenesis 72: A [4-vinyl] chlorophyllide a reductase assay using divinyl chlorophyllide a as an exogenous substrate. Anal. Biochem. 231: 164-169.
- Ellsworth, R. K., and A. S. Hsing, (1974). Activity and some properties of Mg-4-ethyl-(4-desvinyl)-protoporphyrin IX monomethyl ester:NAD+ oxidoreductase in crude homogenates from etiolated wheat seedlings. Photosynthetica. 8: 228-234.
- Whyte, B. J., and W. T. Griffiths, (1993). 8-Vinyl reduction and chlorophyll a biosynthesis in higher plants. Biochem. J. 291: 939-944.
- Suzuki, J. Y., and C. E. Bauer, (1995). Altered monovinyl and divinyl protochlorophyllide pools in bchJ mutants of Rhodobacter capsulatus. Possible monovinyl substrate discrimination of light-independent protochlorophyllide reductase. J. Biol. Chem. 270: 3732-3740.
- Belanger, F. C., and C. A. Rebeiz, (1982). Chloroplast Biogenesis: Detection of monovinyl magnesium protoporphyrin monoester and other monovinyl magnesium porphyrins in higher plants. J. Biol. Chem. 257: 1360-1371.
- Armstrong, G. A., S. Runge, G. Frick, U. Sperlink, and K. Apel, (1995). Identification of NADPH:protochlorphyllide oxidoreductases A and B: A branched pathway for light-dependent chlorophyll biosynthesis in Arabidopsis thaliana. Plant Physiol. 108: 1505-1517.
- Holtorf, H., R. S. Reinbothe, C. Reinbothe, B. Bereza, and K. Apel (1995). Two routes of chlorophyllide synthesis that are differentially regulated by light in barley (Hordeum vulgare L.) Proc. Natl. Acad. Sci.USA. 92: 3254-3258.
- Carey, E. E., and C. A. Rebeiz, (1985). Chloroplast Biogenesis 49. Differences among angiosperms in the biosynthesis and accumulation of monovinyl and divinyl protochlorophyllide during greening. Plant Physiol. 79: 1-6.
- Ionannides, I. M., D. A. Fasoula, K. R. Robertson, and C. A. Rebeiz, (1994). An evolutionary study of chlorophyll biosynthetic heterogeneity in green plants. Biochem. Sys. Ecol. 22: 211-220.
- Shioi, Y., and K. I. Takamiya, (1992). Monovinyl and divinyl protochlorophyllide pools in etiolated tissues of higher plants. Plant Physiol. 100:1291-1295.
- Abd El Mageed, El Sahhar, K. F., Robertson, K. R., Parham, R. and Rebeiz, C. A. (1997). Chloroplast Biogenesis 77: Two novel monovinyl and divinyl light-dark greening groups of plants and their relationship to the chlorophyll a biosynthetic heterogeneity of green plants. Photochem. Photobiol. 66: 89-96.
- Fasoula, D.A., C. Smyth, and C.A. Rebeiz (1996). Relationship of the monovinyl protochlorophyllide a content to plant yield. In: Handbook of Photosynthesis, (M. Pessarakli, ed.) CRC press: Boca Raton, Florida. p. 671-679.
Go Back to Main Menu