![]()
Corraloid Roots of Cycads
by Eric THOUMIRE
St. Maur des Fosses, France
PHYSIOLOGY OF THE
CYANOBACTERIA
When a cyanobacterium has an effective symbiotic relationship with a cycad, it is called a
cyanobiont. Cyanobionts are not specific to the cycads; they have symbiotic relationships
with other plants. May some species be specific to some cycads? It would seem to be true,
perhaps due to chemical signals.
The cyanobacteria have the unusual ability to form chains (filaments) of photosynthetic cells. All those that can become cyanobionts have the characteristic of forming:
Hormogonia are elongated, thin and mobile filaments, which are a kind of transcient escape unit to reach the best condition of living. Hormogonia are induced by red light (this is yet to be confirmed), auxine, chemical mediator of the host, changing of enlightenment, lack of nutrients.
Heterocysts are specialized, oval-shaped cells that are larger than the other cells in a filament; they are able to fix nitrogen and are not movable. They look like this (fifth position): ooooOoooooo and they are permanent. They are designed to protect denitrogenase (an enzyme needed to fix N2) from O2. However, they are not totally closed, for they are able to communicate with their lateral vegetative neighbors in an incredible cooperation totally unusual for bacteria. Heterocysts are induced by the lack of nitrogen.
| CO2 ->-.
.-<----------------LIGHT----------------->-. .-<- N2 _______|_|______ _________|_|_______ | | photosynthesized | | |cyanobacterium-+---> elements -----+--> cyanobacterium | |vegetative cell| + glutamate | heterocyst | | | | | | <-+-- glutamine ---------<--+--- | |_______________| |___________________| |
Necridia are deteriorated and dying cells permitting the scission of a filament.
Akinetes are spores, but there is no evidence that this form occurs in cycad roots. FYI, akinetes of Anabaena have been able to live again after 60 years and Nostoc akinetes have survived 87 years in a herbarium! They are induced by cold, drought, lack of phosphate, and low light with lack of nutrients.
The cyanobacteria do not have the same behavior when they are symbionts compared to the forms that are free-living. For example, heterocysts represent 5-10% of the filament when out of the cycads and 17-45% (and occasionally up to 80%) when inside the cycads. The free-living forms of cyanobacteria are mainly phototrophic ("fed" by light like green plants) and do not require any vitamins to live. They have the same photosystem (phosynthesis system) as the higher plants, but they lack chlorophyll b like in the red algae. They have protein antennae called phycobilisomes made with at least three different protein pigments to catch the light. This equipment provides light absorption throughout the spectrum, from red (565 nm), with a 100% effectiveness, to green (450-650 nm), which cannot be utilized by plants themselves.
The phycobilisomes (approximately 60% of the proteins) can also be considered as a carbon and nitrogen reserve that might be used during prolonged darkness. High luminosity inhibits the synthesis of the phycobilisomes. In high light and the absence of a nitrogen source, the cyanobacteria need no more phycobilisomes and they reuse the nitrogen of the phycobilisomes through their proteolysis. Conversely, the synthesis of the phycobilisomes can be induced with nitrate and lower lights. Additionally, the bacteria can adapt its pigments to the received light.
The symbiont form in cycads
As noted by Norstog & Nicholls, the symbionts become heterotrophic, taking their
carbohydrates from the cycads, via the "transfer cells" (see below). Multiple
heterocysts in each filament are so numerous in cycads that they are supposed to be
ineffective since they do not have enough neighboring vegetative cells to work with. The
nitrogen fixing is less productive than for the free-living form. According to my
readings, phosphorus could be limiting for the cyanobacteria.
The schema is similar, except that the cyanobacteria vegetative cells are replaced by the host cells, as follows:
| CO2 ->-.
.-<---------------LIGHT------------------>-. .-<- N2 _______|_|_______ _________|_|_______ | | photosynthesized | | | HOST -+---> elements -----+--> cyanobacterium | | CYCAD | + glutamate | heterocyst | | | | | | <-+-- glutamine --------<--+--- | |________________| |___________________| | °->- O2 |
CYANOBACTERIA EVERYWHERE
Cyanobacteria exist almost everywhere and have been around since the emergence of
life on Earth. Some emerging theories argue that they (or their ancestors) might have
merged their genotype with those of multicellular beings to be genetically produced by
them the same way our cells can produce viruses. Then they should have become like the
chloroplasts in plant cells (and maybe mitochondria in animal cells.) Well, let's return
now to our less polemic and more accessible subject.
You can find cyanobacteria in/on the ground, on calcareous cliffs, in fresh water (polluted water, you know, I mean 'by lack of oxygen' <"eutrophisation" in French>), in sea water (the name of the Red Sea comes from the red color given off by some cyanobacteria at times), in salt (the salt can be removed with a rake thanks to a layer of cyanobacteria which separates the clean white salt from the muddy bottom), in glasshouses (on glass or wet soils) and many other areas--even on the fur of sloths, where it seems they would be responsible for their blue-grey color! They are missing only where pH < 5.
NAMES OF CYANOBACTERIA IN CYCADS
Cycanobacteria which have "symbiotic" relationships with cycads are in the Order
Nostocales (one of five orders of cyanobacteria). Examples of genera in the Order
Nostocales are as follows: Nostoc, Spirulina, Oscillatoria, Anabaena,
Rivularia, and Calothrix. Nostoc and Anabaena are those more
commonly found in the coralloid roots of cycads, but other Nostocales can also be found,
provided they have the necessary characteristics to infect the roots and to develop and
maintain a symbiotic relationship with the cycad. Species such as Anabaena cycadeae,
Anabaena circinalis, and some others like Nostoc ellipsosporum (in Cycas)
or Nostoc sp. (in Macrozamia lucida), can and do act as cyanobionts.
Additionally, members of the genus Calothrix can infect the coralloid roots of Encephalartos
species. Several genera or species can infect the same cycad at the same time, but one of
them tends to predominate and supplant the others. So, knowing which species is for which
cycad is not really all that important (except for those of you who would like to breed
some from an identified culture from a university).
David L. Jones in "Cycads of the World" reports the following list of cyanobionts:
Norstog & Nicholls, in "The biology of the cycads," also report the following names: Anabaena macrozamiae, Nostoc cycadae, Nostoc punctiformae, Nostoc commune.
GROWTH OF CORALLOID ROOTS
At first, the cycad sprouts without any cyanobacterium in it since the cyanobacteria are
an exogenous contribution of Nature. Corralloid roots are formed soon after the primary
root. I have some Zamia seeds sprouted and I have seen, for each of them, that two
small roots go up after the main root goes down.
 |
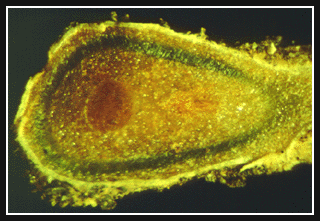 |
| Coralloid roots from the base of a cycad (Dioon spinulosum). The brittle roots contain colonies of nitrogen-fixing cyanobacteria. | Microscopic cross-section of a coralloid root from the base of a cycad (Dioon spinulosum). Just inside the outer cortex is a greenish layer of nitrogen- fixing cyanobacteria. |
| Photos and text descriptions courtesy and copyright © 1999, Wayne P. Armstrong, Wayne's Word; used with permission. | |
Norstog & Nicholls reported the experiences of DeLuca & Sabato (1980) and DeLuca et al. (1980) as follows: "in Cycas revoluta and Macrozamia communis, nodulated apogeotrophic roots will develop on pieces of the gametophyte itself when this is cultured in appropriate conditions in a sterile medium. In this case, the formation of the nodulated roots takes several months and occurs only if the growth factors kinetin (6-furfurylaminopurine) and 2,4-D (2,4-dichlorophenoxyacetic acid) are added to the medium." The trail of a hormonal cause for the development of the coral roots would resolve the problem of the gravity influence.
But what happens in the epiphytic Zamias? Do they sit on a high branch with corraloid roots going upwards? Norstog & Nicholls report also that "it has been suggested that the original (and perhaps continuing) main functions of apogeotrophic roots was as 'pneumatophores'"; further, they write that it would be interesting to see if the coral roots are more developed in the cycads which grow in thick wet mud like Zamia roezlii. All of us know that the cycads generally need a well-drained soil. Can anyone answer about Zamia roezlii or other ones in thick wet mud?
The coral roots may be produced for two reasons:
1) for cyanobacteria but sometimes they have no cyanobacteria. Why?
a) Is there no cyanobacterium around? This is improbable and my friend
told me it is not always possible to inoculate the coral roots even though he brings much
cyanobacteria.
b) The cycads need cyanobacteria but they can't enter the roots? First,
a personal remark: If the coral roots are made by the cycads to be absolutely invaded by
cyanobacteria, it is really strange that, with such an ancient plant and an ancient
bacterium, that the all-knowing Nature forgot to place a mechanism to permit the entrance
of the cyanobacteria. In fact, Nathanielsz & Staff (1975), Staff & Ahern (1993),
and Ahern & Staff (1994) demonstrated that finally the cyanobacteria enter the roots
through wounds. But the cause of the wounds is unknown. The experiment was conducted with
young seedings invaded under controlled conditions. So the cause may come from:
i) the cyanobacteria themselves? Maybe thanks
to the thin and tender periderm of the young roots.
ii) the roots were wounded during their growth
against substrate? Maybe in Nature, but in such an experiment the substrate should be
soft.
iii) The roots were wounded while the seedling
was collected? Why not.
c) The cycads don't need cyanobacteria, so the cyanobacteria can't
enter the roots? Perhaps these plants can decide to let them grow in them or not!?
d) The cyanobacteria can enter the roots but can't develop themselves?
Can the cycad decide to let them grow or not!?
2) There is another use for corraloid roots, other than for cyanobacteria. Perhaps, as we have previously seen, the coral roots are produced to breathe like pneumatophores?
Until now, cyanobacteria show us a kind of opportunism from the cyanobacteria toward the cycads and a possible chemical "password" to recognize the compatible strains. None of them (cycads and cyanobacteria) need the other to live. I write this, although the epiphytic Zamia pseudoparasitica has become epiphytic thanks to their very big corraloid roots (and their very mucilagenous and sticky seeds).
INOCULATION OF CYANOBACTERIA
The entrance of the cyanobacteria
I touched on this point previously: the cyanobacteria need their mobile hormogonia form to
enter the roots and to proceed into them.
They proceed into the
cyanobacterial zone
They now have to join the cortical cyanobacterial zone (inaccurately called 'algal zone')
situated in the cortex (under the periderm), where they can become a cyanobiont. To do
this, they must cross between or though cells and
| 1 2 3 4 5 4 3 2 1 || |=| | | |=| || 1=periderm || |=| | | |=| || 2=outer cortex (this zone does not exist in || |=| | | |=| || Norstog & Nicholls' schema but is quoted by || |=| | | |=| || other sources) || |=| | | |=| || 3=cyanobacterial zone || |=| | | |=| || 4=inner cortex \\ \ \ \ / / / // 5=stele \\ \ \ \/ / / // |
Setup of a symbiotic relationship
In response to nitrogen deprivation (Bergman, Matveyev & Rasmussen, 1996),
cyanobacteria become the fixed heterocyst form, fixing nitrogen. So we can say that a
cyanobiont IS a heterocystous filament.
DISPLACEMENT AND GROWTH OF
CYANOBACTERIA IN ROOTS
Cyanobionts are known to be very slow growing, at least in other plants (Gunera, in
Söderbäck & Bergman, 1992). If this is also true for the cyanobacteria in cycads,
they might take a long time to become effective if used to rescue a deficient cycad.
Fertilizers would activate much quicker.
The cyanobiont can create the hormogonia form to move into the roots to inhabit better areas for their growth according to their conditions of living or/and chemical signaling. According to M. C. Ow, M. Gantar & J. Elhai (unpublished), "[c]ycad root extracts can also promote hormogonia formation by several thousand fold in competent Nostoc strains."
Norstog & Nicholls report that in deep roots (obscure area) the cyanobacteria seem to become heterotrophic, taking nutrients from the cycad (since there is no other explanation I guess). With the same idea, Bergman, Matveyev & Rasmussen (1996) supposed they might use the mucilage as a source of carbon. Norstog & Nicholls report that Caiola (1980) stated the following: "The cyanobionts, moreover, can remain in an unchanged state in the roots" (as if the cyanobacteria need no more growth or the cycad need no more nitrogen?) "even while the cycad is seasonally inactive in terms of its general metabolism and growth." Personally I would rather say "especially while..."
NITROGEN FIXATION BY CYANOBACTERIA
Denitrogenase is an enzyme containing molybdenum (Mo), so Mo is needed to fix N2.
With a lack of Mo and N2, Anabaena cylindrica produces more heterocysts
(to fix N2) but its nitrogen fixation decreases (heterocysts ineffective by
lack of Mo).
About the moving of the cyanobacteria downward into the roots (and the dark) which might induce nitrogen fixing, my friend explained to me that the contrary (too much light) causes a proteolysis of the phycobilisomes and consequently the shutdown of the nitrogen fixation by lack of phycobilisomes. But I don't know if such a thing may occur often on non-free-living cyanobacteria. Norstog & Nicholls report that this moving downward into the dark may induce nitrogen fixing. However, these authors quote Bergersen et al. (1965), who demonstrated that "in darkness, the nitrogen-fixation rate was reduced to less than half of this value." Can anyone explain this?
Halliday & Pate (1976), also showed that, for Macrozamia riedlei, the rates are lower at night. And also that "in controlled experiments, nitrogen fixation was fairly constant over the range 15-30°C" but outside they registered seasonal variations at these temperatures and they think there was a rainfall effect (highest rates in winter, lowest rates in summer in the Mediterranean climate of southwestern Australia).
EFFECTIVENESS OF NITROGEN FIXATION
David L. Jones (Cycads of the World) reported that Macrozamia riedlei is
fixing between 19 and 35 Kg N2 /ha/year. As for Norstog & Nicholls, they
report [Halliday & Pate (1976)] that the roots of Macrozamia riedlei "fix
about 37.6 g of nitrogen per kilogram of root tissue per year. Assuming that this is
passed on to the cycad plants, the time taken to double the whole plant nitrogen by the
process of nitrogen fixation would be in the range of 8-11 years". How much time does
it take to double the whole plant nitrogen by the use of fertilizers? I guess it depends
on the plant size. Halliday & Pate indicate 500 kg of cycads per hectare for this
experiment. But I don't think it is enough to calculate an accurate figure.
You can compare these figures with those of the leguminous plants fixing N2 (100-200 kg N2/ha/year) but many small plants is not the same as a few big cycads (not the same physiology). Thus, it is difficult to evaluate. Just FYI, the free-living Anabaena can produce 2 kg N2/ha/DAY (!) in polluted waters.
To explain the ineffectiveness of the cyanobacteria that happens sometimes in the coral root, Norstog & Nicholls report that "recent work, admittedly on very few species, has demonstrated convincingly that nitrogen is fixed in substancial amonts by the cyanobionts in situ, that the nitrogen is made available to the host, and that it is then eventually returned to the environment."(!) I think this tends to show that nitrogen fixing could be induced by another (external) factor other than a cycad demand.
BREEDING OF CYANOBACTERIA
Norstog & Nicholls report that cyanobacteria taken from coral roots can be grown on
cotton (with light of course) and that they can, once again, fix nitrogen. It would be
interesting to keep all the unused cut roots of healthy cycads to put them on the ground
in order to help further inoculations.
PRACTICAL APPLICATIONS
I must admit that the below results reflect the views of people like Tom Broome, who
deduced them months ago. Accordingly, my thoughts are welcome to critique. Here is what I
think:
CONCLUSION
Cyanobacteria are not the panacea for a grower but can save the lives of some cycads. Of
course, I have more questions, other than those above, left without answers. But I will
finish on this subject for now. Below are sites where you can find information about the
cyanobacteria, with details that I have avoided, so as to be not too soporific.
http://chemweb.urich.edu/~sjanson/disp.htm#Nitrogen_fixation
http://WWW-Cyanosite.Bio.Purdue.Edu
http://chemweb.urich.edu/~biology/cyanonews
Another source of helpful information could come from this research below:
Zimmerman, W. J., & B. H. Rosen. 1992. Cyanobiont diversity within and among cycads of one field site. Canadian Journal of Microbiology 38(12):1324-1328.
BIBLIOGRAPHY
Book "The biology of the cycad", by Knut J. Norstog and Trevor J. Nicholls,
Cornell University Press
Book "Cycads of the world", by David L. Jones, Reed Ed.
M-A.Selosse. 1996. Les cyanobacteries, d'etonnants procaryotes autotrophes. Biologie
Geologie (3) 481-529. (in French)
Article "A mode of entry of blue-green algae into the apogeotropic roots of Macrozamia
communis."
by Charmian P. Nathanielsz and Ian A. Staff in American Journal of Botany 62(3):
p 232 to 235. - 1975
Article "Chemical signalling in cyanobacterial-plant symbioses" by Birgitta
Bergman, Andrey Matveyev and Ulla Rasmussen in Trends In Plant Science - Vol. 1
No. 6 - p 191 to 197 - June 1996
![]()
| PACSOF Home Page | |||
| Virtual
Cycad Encyclopedia Site Map Powered by FreeFind. |
|||
This site is copyrighted © 1998, 1999, 2000, Palm & Cycad Societies of
Florida (PACSOF)
For questions or comments, e-mail the webmaster.
Internet hosting provided by Zone 10,
Inc.