Botany for the Next Millennium:
I. THE INTELLECTUAL: EVOLUTION, DEVELOPMENT, ECOSYSTEMS
The tree is an aerial garden, a botanical migration from the sea, from those earlier plants, the seaweeds; it has a purchase on crumbled rock, on ground. The human, standing is only a different upsweep and articulation of cells. How treelike we are, how human the tree. -Gretel Ehrlich2
[Evolution and Diversity] | [Development and the Organism] | [Ecosystem Structure and Function]
Despite the current social and political questions over the relevance of basic science to late 20th century life, the aim of most scientists remains a more complete understanding of how the world works. Plants, with their physically and functionally dominant place in the world, are key to that understanding. They also happen to be inexpensive and convenient as research subjects, without many of the ethical and anthropomorphic issues involved in anima) research. With few exceptions, any aspect of basic cell biology can be effectively investigated in plants. In addition, plants have evolved a suite of unique and important biological characteristics that are found in no other life forms and are of particular interest to science.
EVOLUTION AND DIVERSITY
In the past five to 10 years, the bioscience literature has become filled with family trees-stick diagrams depicting the evolutionary relationships among a group of organisms and the sequence of speciation events that produced them. This explosive area of phylogenetic analysis, helped along by the ability to examine and compare sequences of DNA, addresses the overarching theme in biology: the evolution of ourselves and the millions of other distinct species. As an evolutionary group, green plants are distinctive for their boggling array of ways to reproduce but singular way of feeding-through photosynthesis-that predominates in the biosphere. Our understanding of the history of life on Earth, therefore, is incomplete until we understand the evolutionary processes that have generated the great diversity of plants.
'An abominable mystery" is how Darwin described what seemed to be the sudden appearance of flowering plants in the fossil record. For the last 100 million years they have been the most common type of plant on the planet, outdistancing the ferns and other early types that dominated in previous eras (so abundant were these earlier types that today we extract their decayed and compressed remains as coal and other fossil carbon fuels.) The mass of just one kind of flowering plant, the woody species, today exceeds that of all other kinds of organisms. What prompted the evolution of the elaborate reproductive structures we call flowers, with such prolific and diverse consequences?
A significant understanding of this question has emerged in recent years, even if Darwin's mystery is not yet solved. The presence of chloroplasts in plant cells has given plant scientists a useful tool for comparing the evolutionary changes across groups of plants, since chloroplasts are the site of photosynthesis and contain a piece of DNA that is abundant in the cell. Such analyses, along with studies of the morphological and anatomical differences among plants, have spurred substantial progress in the reconstruction of plant ancestry (it has been said that the plant sciences are far ahead of the animal sciences in this area). As the shape of this major branch of evolution is realized, the forces that make it so can be better understood: how genetic changes conspire with the environment to create new species.
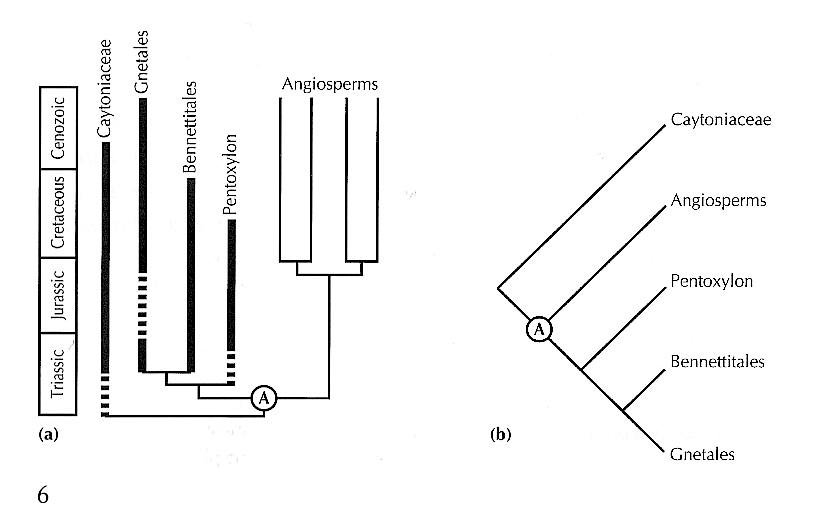 |
|---|
A phylogenetic tree (a) and cladogram (b) for higher seed plants (after James A. Doyle and Michael J. Donoghue. 1993. Paleobiology 19: 141-167.), indicating that the origin of angiosperms must be as old as the late Triassic. Redrawn from: Andrew B. Smith. 1994. Systematics and the Fossil Record: Documenting Evolutionary Patterns. Blackwell Scientific Publications, Oxford. |
The process of these studies is something like map making: unknown areas become pieced together in a way that allows a more meaningful exploration of any one part. One part of the map that has recently been thrown into relief is the assortment of plants that produce mustard oils. Fifteen different families of plants-everything from Brassica, the source of the yellow seed for hot dog mustard, to capers (Capparis), to papayas (Carica), to Drypetes, a poinsettia relative-produce the pungent, insect-deterring chemicals. Given the diversity of the plant forms, most opinion held that the plants were not closely related, and that their shared capability for producing mustard oils was coincidence, evolving independently several times over. But analysis of the chloroplast DNA supports the radical view that most of the mustard oil plants are closely related, except for Drypefes. The distinctive chemistry, therefore, evolved only twice. The finding gives an especially useful frame of reference for the large body of work on the experimental plant Arabidopsis a member of the mustard family.
Although our knowledge of the ancestry of plants has been greatly bolstered by the data flowing from analyses of DNA, only a small number of gene sequences have been examined. There is a vital need to expand that number and integrate the evolutionary history the sequences tell. Still, understanding the process of evolution and diversity will depend on more than these data. As the National Research Council's Committee on Research Opportunities in Biology stressed for evolutionary biology, "advances come from the synthesis of previously disconnected areas, from new ways of looking at problems, or from new concepts."
The flowering plants attract most of botanists attention. Other plant groups are much
 |
|---|
| Choleochaete represents a modern group of green algae (the Charophyceae) whose ancestors were closely related to the earliest land plants. Because the fossil record of the algae-plant transition is sparse, comparative study of modern charophytes and simple seedless plants can illuminate the evolutionary process that generated a terrestrial flora. Courtesy: Linda E. Graham. |
older evolutionarily, simpler to study, and represent the genetic storehouse from which our trees, lawns, crops, and decorative plants drew. These older plant groups live on today as mosses, liverworts, hornworts, clubmosses, horsetails, and ferns. All pose questions in evolution and other areas that have scarcely been addressed.
Algae, commonly thought of as green pond scum, have been found to be extremely diverse: they contain at least seven distinct ancient evolutionary lineages, one of which gave rise to the plant kingdom. As such, the algae represent a myriad of experiments in adaptation, amenable to comparative studies because of their diversity and relative simplicity.
One group of today's green pondwater denizens (Coleochaete) resembles the ancient plants that were the first to grow on land around 440 million years ago. But what accounted for this pioneering transition. We still do not understand the origin of land plants and their early diversification very well. Rocks from this very early period during the Paleozoic have scarcely been examined for plant (spore) fossils. The data on ancient plant diversity from fossil plant records that do exist need integration with current patterns of diversity to improve the picture of life's evolution.
Another pool of ancient, little-known plant diversity exists in the mosses and their relatives, collectively known as bryophytes. Most of these are the small, inconspicuous plants we may barely notice as green carpeting underfoot, on rocks, or on trees. Yet they are significant components of the natural world, abundant in most woodlands, fields, and even urban parks, and the dominant vegetation in tundra and boreal regions. Once considered to have been static evolutionary dead ends, they contain a high degree of genetic variability, a virtually untapped source of information.
These ancient plants, and their descendants today, lived most of their lives with but a single set of chromosomes, a state known as haploidy. The general trend in evolution has been away from haploidy, toward diploidy (double the chromosomes) and the greater diversity of genotypes it makes possible. This very trend can be observed and studied in plants in a way not possible with other organisms. Ferns, as an example, go through both modes as separate plants during their life cycles: whole plants are genetically all haploid or all diploid.
Many plants can even be polyploid, with multiple sets of chromosomes. Ferns appear to have taken this phenomenon to the limit: Ophioglossum has about 1,260 chromosomes, thought to represent at least 10 full sets. Evolution through polyploidy appears to have been very important in many lineages of plants, whereas this evolutionary door seems all but closed to animals. In humans, individuals with even a part of one chromosome either missing or replicated often have significant abnormalities.
How the size and other physical aspects of chromosomes affect evolution is one of the current evolutionary questions plants are particularly suited to answering. Many other such questions are being asked, or deserve to be asked. A few examples:
- the coevolution of plants and other organisms, including the intriguing process that created lichens
- the association of a fungus and a photosynthesizing cell
- the evolution of the developmental pathways and diverse mating systems in plants
- the role of disease in population changes and hence in evolution
- the differences in photosynthetic apparatus across several evolutionary lines
DEVELOPMENT AND THE ORGANISM
The question of how the complexity of an organism develops from the simplicity of a single cell remains one of biology's enduring mysteries. In plants, the question is all the more striking because of their clonal powers: a piece of a plant, even a single cell, can regenerate a genetically identical plant. It's a familiar concept to anyone who has pulled a dandelion without getting the whole root. Similarly, a patch of clover can be the clonal growth of a single plant that has propagated by sending out runners that send up identical shoots, which in turn send out new runners.
We don't know why plants have this potential and animals generally do not; somehow,
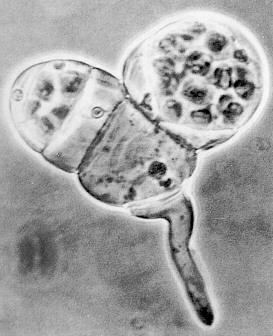 |
|---|
| Gametophyte of the fern Bommeria hispida. The gametophyte consists of a single rhizoid, one gametophytic cell, and two antheridia. From: Christopher H. Haufler and Christopher B. Welling. 1994. American Journal of Botany 81: 616-621. |
plant cells retain an embryonic quality (totipotency) that most animal cells lose. As a result, you can buy a Boston fern produced by cloning, but not a Boston terrier. The technique has been put to use in biotechnology, as well as in the study of development, but is little appreciated outside of plant biology.
Almost ironically, even though most cells in a plant have this regenerative power, growth in a plant is the specialized purview of only a few cells, collectively known as meristems. When trees leaf out in spring, it is the result of division and differentiation of meristematic cells in the shoots. In many ways the perpetually young meristem offers a distinct system for studying development: all of a plant's future parts its leaves, stems, flowers, fruits eventually develop from these few cells. The process lends itself especially well to study in the relatively simplified meristems of certain ferns.
All the physical and biochemical signals that sculpt an organism's development must somehow be read by the meristem. Sunlight, a few hormones (with myriad functions), and other factors such as nutrient levels contribute to the critical switch by the meristem from simple vegetative growth, characteristic of a leafy shoot, to reproductive growth, such as the development of a flower. Or the plant will differentiate into a male or female, another intriguing area of investigation. In some ferns, for example, male haploid plants do not appear until one vigorous specimen secretes a hormone that induces the surrounding plants to become male. Understanding how such switches work, specifically how genes are tripped and lead to the physical alteration of growth, will enhance our knowledge of development in genera.
Plant growth is also distinctive for its plasticity, a poorly understood phenomenon. Perhaps in part because they are rooted, land plants have an ability to modify their growth habit to an astonishing degree. Plants cannot move to escape a predator, competitor, or harsh environmental conditions. Instead, they respond by modifying their growth and development to yield altered structures. Even clones of the same individual plant can appear to be confoundingly different depending on the conditions they are grown in. Shade-covered suckers growing at the base of a tree generally have larger leaves than those at the sun-lit tree top, even though they are genetically identical. Similarly, in ferns and many other plant groups, diploid plants are strongly differentiated from their haploid forms, while in some groups of algae, for example, haploids and diploids are virtually indistinguishable. In effect, the dramatic changes (or not) in plant appearance address the larger question of nature versus nurture, how genes and the environment they find themselves in play out in the organism.
The striking aspects of plant development are worthy of understanding in their own right. Yet many basic mechanisms common to all organisms are accessible for study in plants as well. Some fungi, notably budding yeast, have been and continue to be models for explaining basic genetic aspects of development. They will no doubt help illuminate the crucial moment when a dividing cell begets a cell that is in some subtle way different from its sister, the origin of differentiation. Algae and mosses (bryophytes), as well as ferns, also offer systems of differentiation that are simpler and easier to study than in those of seed plants and animals. In a haploid bryophyte, for example, the effect of all its genes can theoretically be studied, since recessive genes are not masked as they are in the diploid state.
New imaging techniques are allowing researchers to visualize physical changes in cells as the cells differentiate. Such approaches offer the promise of joining the wealth of molecular data that has accumulated for plant cells to a picture of what is happening to the organism. Technical breakthroughs of the last 20 years have allowed our biochemical knowledge of plants to race far ahead of organismal and ecological knowledge. The next challenge in development is to integrate vertically the molecular data with effects on the organism, and then the ecosystem. This work has now begun. In maize, for example, a mutation that turns the normally male part of the plant (the pollen-producing tassel) into the female part (the ear) has been traced to a hormone-modifying enzyme.
Not every molecular aspect of plant function is understood, however, including photosynthesis. Melvin Calvin received the Nobel prize in 1961 for working out, in algae, how plants absorb and transform carbon dioxide into sugar, but major twists on the basic process have been discovered as recently as the 1970s. Today, investigation centers on the complicated structure involved in splitting apart water and generating oxygen, the remaining black box in the process, as well as how photosynthesis is affected by rising carbon dioxide levels and increased ultraviolet radiation (see below). The key enzyme involved in fixing carbon dioxide, ribulose-1,5-bisphosphate carboxylase/oxygenase, is an important target of study. It's an interesting enzyme for its sheer quantity3; its seeming inefficiency in that it can bind both carbon dioxide and oxygen; its relatively novel construction (with the help of molecular chaperones) from amino acids encoded within the plant's chloroplast DNA as well as the nuclear DNA; and the slow rate of mutation of the chloroplast sequence, which has made it instrumental in tracing the evolution of photosynthesizing organisms.
ECOSYSTEM STRUCTURE AND FUNCTION
We naturally think of land in terms of the plants it harbors: grassland, cropland, woodland, forestland. The tropics conjure jungle; the desert suggests cactus. These names and images accurately reflect how plants define places not only structurally but functionally, in producing the energy and food upon which animals depend. Their defining role warrants special attention to plants if we are to understand how ecosystems work, especially under today's unprecedented conditions of human-influenced environmental change.
We know basically how plants are built and how they can create structures as massive as a 10-story tree: dividing plant cells don't move as animal cells do, but are laid down like bricks, encased in a fibrous wall of cellulose, providing with lignin great mechanical strength and rigidity. But why plants are built the way they are and the consequences, if any, for the rest of the community of a plant's leafing pattern, stem height, or other physical qualities are only beginning to be investigated. Traditionally, botanical texts have described the differences in plant structures among different species, a wealth of diversity apparent to the most casual observer. But ecological or evolutionary discussions of the differences have been missing, except for certain striking forms such as cacti, which substitute spines for leaves, store water in enlarged stems, and take in carbon dioxide mainly at night. The result is a heat-dissipating, desiccation-resistant desert plant that tastes sour at night but sweet by the end of the day, when the store of carbon fixed during the night is turned into sugar.
An ecological understanding of plant form and function is especially lacking for the tropics, where plants display their greatest diversity in form. Most of our knowledge of plants is based on studies of temperate, often agronomic, plants. The structural innovations of plants in frost-free environments, which may differ dramatically from familiar temperate representatives, are too often characterized as exceptions or anomalies, rather than examples of the full potential of plant growth. Palms and bamboo, for example, which are in the same class (monocots) as grasses, are woody and treelike, and trees in the tropics lack the familiar rings of annual growth. Many fascinating structural modifications in tropical plants are related to complex interactions (associations) with
 |
|---|
| Jacana (Bediparra gallinacea) on a lily pad; Kakadu, Australia. Courtesy: Ray F. Evert. |
animals: some tropical plants (melastomes) have little swellings on their leaves that serve as housing for ants. The developmental basis of much of this structural and architectural diversity remains to be studied.
Functionally; plants are the primary mediators between the physical and biological world. Energy, carbon, hydrogen, oxygen, and nitrogen, the most basic ingredients for life, all enter the biological world primarily through plants. Fungi and bacteria, by digesting the plant and animal products of photosynthesis, return the elements to their inorganic form. Green plants also exert a heavy influence on the cycling of water. The loss of water from plants by transpiration returns enough vapor to the air to account for half of all rainfall by some estimates.
Although we have identified where and how plants fit into these global cycles, we fall short of understanding ecosystem effects of perturbations on the cycles. The problem of scaling up what happens in a leaf to a whole plant to a community to a biome and the planet remains a challenge. And we are still uncovering the ways plants affect their communities and large-scale cycles. For example:
Mosses, other bryophytes, and lichens play little understood but significant roles. They sponge up nutrients and take toxic metals out of the system. The sphagnum (peat) mosses are of immense importance in the global cycling of carbon-they may cover as much as 2 percent of the Earth's surface.
Certain tree species emit large amounts of hydrocarbon gas with as yet unknown natural effects, but with potential implications for air quality policies.
The bulk of photosynthetic production in the oceans turns out to be the work of particularly tiny algae known as picoplankton, and their productivity is significantly affected by the availability of iron, a simple and surprising phenomenon. Moreover, a delineation of plankton growth rates and distribution remains elusive, one of the stumbling blocks for understanding global climate change.
The interactions among organisms create an ecosystem and are a rich area of inquiry. Interactions involving animals, especially predation but also pollination and herbivory, have received a great deal of attention from biologists, although plant interactions may be more important on an ecosystem level. Associations between mycorrhizal fungi and plants are turning out to be almost standard-most seed plants have these filamentous organisms growing in and about their roots. Yet the nature of the association isn't definitively known. The mycorrhizae (literally fungal roots) aid the plant in absorbing nutrients-many trees in the nutrient-poor soils of the tropics have them-but it's less clear what the fungi gain, since they are able to live freely in the soils as well.
2Ehrlich, Gretel, "River history," in Montana Spaces: Essays and Photographs in Celebration of Montana, William Kittredge, ed (New York: Nick Lyons Books, 1988, 69-72.
3Because it is so abundant 20 percent of the protein in a leaf is RuBP carboxylase/oxygenase-it has been called the most common and important protein on Earth and touted as the ultimate food; the idea once prompted a joke during a presentation at a meeting involving "rubisco crackers," a play on the trade name Nabisco. Although the snack food suggestion fell flat, the abbreviated name rubisco has stuck in the scientific literature.




