
Botany online 1996-2004. No further update, only historical document of botanical science!
Energy-converting biological processes are usually membrane-bound. In this, they differ fundamentally from simple enzyme reactions happening in solutions. A gain of energy became possible only after membrane-surrounded vesicles, i.e. cells or at least cell precursors had evolved. In the course of the evolution of photosynthesis and the respiratory chain, proton gradients developed. They polarize the membrane, and the energy stored in these gradients is used for the production of an energy-rich bond, a phosphorylation (ADP + Pi > ATP). The cell uses the energy stored in this bond for other processes.
Model experiments show that photophosphorylation, i.e. phosphorylation as a consequence of the use of light energy may also occur under abiotic conditions – though with a small yield only – and in the absence of membranes, but in the presence of molecules that might have existed already shortly after the formation of the earth’s surface (see the simulation experiments of MILLER in the topic ‘How Did Life Evolve?):
haemin + imidazole + Pi + ADP > (Light) > ATP
(W. S. BRINIGAR, D. B. KNAFF, J. H. WANG, 1967).
Primitive procaryotes can cover their energy expenses by using the electrons of inorganic material (chemolitho-autotrophy), more advanced organisms ferment organic material (heterotrophy).
A simple transformation of light energy into chemical energy occurs in the archaebacterium Halobacterium halobium. Just a single protein participates in its ‘photosynthesis’, while the electron transport chain that is typical for more advanced energy-yielding mechanisms is missing. The energy yield of Halobacterium is extraordinarily small.
The photosynthesis that is typical for green plants, blue-green algae, and photosynthetic bacteria is based on the use of light energy (originally with a high portion of UV), i.e. on the conversion of a flow of photons into a flow of electrons and on the existence of electron transport chains.
Photoautotrophic organisms contain usually chlorophyll a or a related derivative like the bacterial chlorophyll a. The porphyrin part of chlorophyll molecules contains magnesia ions instead of ferro- or feriions that are the central atoms in haemin molecules. The change to magnesia occurred most likely, because the new complex made better use of the available amount and quality of the light emitted by the sun.
The membrane proteins bound to chlorophyll a as well as the composition of the other "accessory pigments" differs among species. It is assumed that the primitive atmosphere was reducing. Hydrogen, hydrogen sulfide and other sulfide compounds may have acted as electron donators.
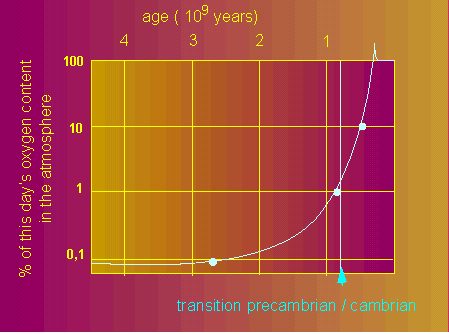 Blue-green algae (Cyanophyceae, Cyanobacteria) were the first organisms to use water as both a proton and an electron donator. As a consequence of their early photosynthetic activity, free oxygen entered the atmosphere and its concentration rose from originally almost zero to roughly 20 percent. This changed the conditions of selection for all organisms considerably. Those able to protect themselves from oxygen and eventually able to use it did finally succeed.
Blue-green algae (Cyanophyceae, Cyanobacteria) were the first organisms to use water as both a proton and an electron donator. As a consequence of their early photosynthetic activity, free oxygen entered the atmosphere and its concentration rose from originally almost zero to roughly 20 percent. This changed the conditions of selection for all organisms considerably. Those able to protect themselves from oxygen and eventually able to use it did finally succeed.
The increase in the amount of oxygen led to the development of an ozone layer in the stratosphere and thus to a shift in the spectral composition of the light reaching the earth surface. Short waves (UV) were now kept back almost completely. The organisms had thus the opportunity to use the visible light increasingly more and as a consequence, the pigment composition of the photosystems changed.
A further understanding of the evolution of energy-converting processes has to consider the evolution of the electron transport chain. Among the similarities of photosynthesis and the respiratory chain is the use of the same proteins: ferredoxin, cytochrome c, cytochrome b, and several others.
The cytochromes b6 (from plastids) and b (from mitochondria) can be homologized due to their topology. Cytochrome b6 corresponds in its change of hydrophobic and hydrophilic amino acid sequences to a sequence at the N-terminus of cytochrome b (W. R. WIDGER, W. A. CRAMER, R. G. HERRMANN, A. TREBST; Purdue University, University of Munich, Germany, University of Bochum, Germany, 1984). The fact that corresponding cytochromes can be homologized is a further proof for a common ancestor of photosynthesis and respiratory chain.
A comparison of the topology of the complexes participating in the electron transport chain leads to a similar conclusion. The respiratory chain is thus a variation of photosynthesis. It was perfected in parallel to the increase of free oxygen.

The photosystems I and II developed at first independently. Combination led to a system, the Z-scheme of photosynthesis, that is far more capable to use light energy efficiently than each of its partial systems alone.
