On Archaeopteryx, Astronomers, and Forgery
Copyright © 1996-1997 by
Chris Nedin
[Last Update: December 15, 1997]
![]()
URL of this page: http://www.talkorigins.org/faqs/archaeopteryx/forgery.html
On Archaeopteryx, Astronomers, and Forgery
Copyright © 1996-1997 by
Chris Nedin
[Last Update: December 15, 1997]
![]()
Contents
 rchaeopteryx
lithographica is regarded as one of the most
important fossils ever discovered. This isn't because of any uniquely
transitional nature, since many transitional forms exist (e.g. the
synapsid to mammal transition), but due to the fact that
Archaeopteryx is such a good example of evolution. The
skeleton is essentially reptilian, with close affinities to theropod
dinosaurs, and possesses teeth, a long bony tail, abdominal ribs and
three digits on each hand - characters absent in birds. However, the
specimens also show certain bird characters such as a furcula
(wishbone) and a retroverted pubis (characters also shared with some
dinosaurs) and a opposable hallux (big toe) for perching. Along with
these other avian characters, the most spectacular feature is the
distinct impression of feathers around the forelimbs and tail,
feathers almost exactly like those of modern birds.
rchaeopteryx
lithographica is regarded as one of the most
important fossils ever discovered. This isn't because of any uniquely
transitional nature, since many transitional forms exist (e.g. the
synapsid to mammal transition), but due to the fact that
Archaeopteryx is such a good example of evolution. The
skeleton is essentially reptilian, with close affinities to theropod
dinosaurs, and possesses teeth, a long bony tail, abdominal ribs and
three digits on each hand - characters absent in birds. However, the
specimens also show certain bird characters such as a furcula
(wishbone) and a retroverted pubis (characters also shared with some
dinosaurs) and a opposable hallux (big toe) for perching. Along with
these other avian characters, the most spectacular feature is the
distinct impression of feathers around the forelimbs and tail,
feathers almost exactly like those of modern birds.
The authenticity of Archaeopteryx, or more specifically the authenticity of the feather impressions, was questioned in 1985 by a group which included, Prof. F. Hoyle (astronomer), Dr. N. Wickramasinghe (mathematician), Dr. L. Spetner (physicist), Dr. R. Watkins (medical doctor) and J. Watkins (photographer) in a series of articles published in the British Journal of Photography (Hoyle et al. 1985; Watkins et al. 1985a, 1985b, 1985c). Interestingly, one of the authors (Dr. Spetner) was claiming that Archaeopteryx was a fake as early as 1980 (Trop 1983). Apparently on the sole fact that the London specimen was sold by Dr.C Haberlein and the Berlin specimen was sold by Dr Haberlein's son! (The Harberleins were well known collectors, possessing one of the finest collections of Solnhofen fossils).
Needless to say, these claims were vigourously opposed by the British Museum of Natural History (BMNH). Several people within the Museum collaborated to refute the claims of forgery. These were A. Charig (chief curator of fossil amphibians, reptiles and birds, BMNH), A. Milner (principle scientific officer - fossil amphibians, reptiles and birds, BMNH), C. Walker (senior scientific officer, fossil amphibians, reptiles and birds, BMNH), F. Greenaway (principle photographer, BMNH) and P. Whybrow (photographer, BMNH).
Watkins et al. Opening Statements
In the first part of their claim that the feather impressions were a forgery, Watkins et al. stated that,
"Although several other reptilian fossils have subsequently been reclassified as Archaeopteryx, the first two referred to above [the London and Berlin specimens - cn] remain unique in that they are clearly of the same prototype and possess unmistakable feather imprints." (Watkins et al. 1985a, p. 264-265).
This was reiterated by Hoyle et al. (1985, p. 694), who suggested that
"the only undeniable featherlike impressions are therefore those on the single feather of 1860, on the British Museum specimen of 1961 and the Berlin specimen of 1877".
This is incorrect. It is true that none of the other specimens have feather impressions as good as those found on the London and Berlin specimens and that the recognition of feathers on the Haarlem specimen would not be possible without reference to the London and Berlin specimens. However, the Eichstatt specimen has clear feather impressions (Wellnhofer 1974) and the Maxberg specimen has impressions in which the structure of the feather is discernable as being typical of that in modern birds (de Beer 1954; von Heller 1959; Charig et al. 1986). Not only that, the feathers of the Maxberg specimen clearly refute any possibility of forgery because they continue under the bones of the skeleton and are overlain by dendrites (von Heller 1959; Charig et al. 1986). (Dendrites are crystal aggregates occurring along flat surfaces, with a tree-like branching pattern. They often occur in cracks or along bedding planes). The Haarlen specimen does show faint feather traces (Ostrom 1972). Feather traces have also been described from the Solnhofen specimen (Wellnhofer 1988). The most recent find, the Solnhofen-Aktien-Verein specimen, has also been described as possessing feather traces (Wellnhofer 1993).
Another statement is,
"The significance of Archaeopteryx lies in the fact that it represents the only unquestionable case of a fossil showing a transition between two vertebrate classes, aves (birds) and reptilia (reptiles)." (Watkins et al. 1985a, p. 256).
Again, this is incorrect. There are numerous examples in the fossil record, with probably the best documented example being the transition between the synapsid reptiles and the mammals (e.g. Kemps 1982; Benton 1990; Colbert & Marales 1991).
A third statement concerns the photography itself. In 1984, Watkins et al. took comprehensive photographs of the London specimen, held at the BMNH, on colour transparency film with a hand-held 35 mm SLR camera and low angle tangential flash lighting. The resulting slides were then enlarged by projecting the slide onto a distant screen and by making black and white prints (Watkins et al. 1985a). When describing the technique used by Watkins et al., they comment,
"Such [photographic] studies have been made earlier but these have inevitably been limited by the techniques available in the past." (Watkins et al. 1985a, p. 265).
This is also incorrect. As the response from the Museum indicates,
"As professional photographers we have studied the fossil under various combinations of light sources, emulsions, ultraviolet reflectance and fluorescence, filtered UV fluorescence, intensive scanning by infrared TV and high photomicrography. These studies have taken place over a number of years." (Parmeter & Greenaway 1985, p. 458).
Indeed, far from being in any way superior to other methods,
"the cursory examination and poor photographs of the authors of the articles [Watkins et al. - cn] bear no comparison with the close scrutiny and exacting standards of the Museum." (Parmeter & Greenaway 1985, p. 458).
The photographs themselves have too much contrast and too soft a focus (Charig et al. 1986), making detailed study difficult and these
"newer photographs compare extremely unfavourably with photographs of the same specimen taken by museum photographers who, several decades ago, were already using low-angle oblique lighting with far greater success" (Charig et al. 1986. p. 623) (See de Beer 1954 for an example of this.).
Indeed, Watkins et al.
"readily concede the rudimentary nature of their photography, but were looking in the time available for evidence to prove or disprove certain specific theories. Ideally they might first have inspected the Museum's photographic records, but, as ever, those holding controversial views preferred to look for themselves before disclosing them" (Crawly 1985, p. 458).
In yet another statement, the authors state,
"It is now generally believed that the skeleton is largely reptilian except for the furcula (wishbone) which is bird-like" (Watkins et al. 1985a, p. 256).
This is misleading. The opposable hallux is also an avian feature as is the position of the pelvis (although this also occurs in some dinosaurs) (see for instance, Ostrom 1976).
The Main Claims and Evidence, For and Against
Using a series of photographs, Watkins et al. claimed that the feather impressions were fakes. The method used to create the forgery was via the pressing of chicken feathers into a thin layer of artificial cement surrounding a small reptile skeleton. (It should be noted here that although one of the authors - Dr. Spetner - claimed that chicken feathers were used, the impressions do not correspond to chicken feathers, but are more like rail feathers). The cement would have been made by mixing limestone from the same deposit with some binder and which was spread thinly over the surface of the slabs. As corroborating evidence of this several observations were cited:
i) The difference between the surface textures of the limestone in the feathered and unfeathered areas was cited as evidence of the presence of a cement layer around the feathers (Watkins et al. 1985a).
The difference in surface texture is certainly real. However Charig et al. (1986) explain it as being due to the impression of the animals body on parts of the surface. An analogy used was likening it to the differences in texture seen between a human footprint in mud and the surrounding mud.
"In other words, it was the feather impressions that caused the differences in surface texture; not that a difference in surface texture (due to some other cause) permitted the preservation of the impressions in some places and prevented it in others" (Charig et al. 1986, p. 623).
If a layer of cement is present, then some sort of discontinuity should be visible between the true limestone and the cement, on the surface and/or in vertical section (a vertical section is a section cut through the slab, at 90 degrees to the fossil). No such discontinuity has been found, even in vertical section. There does appear to be a division in vertical section whereby an upper 500-850 micrometre (1 micrometre = 1/1000 millimetre) layer is separated from the lower layer by a dark band. However, the upper layer shows the same granular structure as the lower layer and the structure is continuous through gaps in the dark band (Charig et al.. 1986). Also the complete lack of air bubbles and the presence of calcite crystals indicate that the whole section is original. Besides, the upper layer is far too thin to receive any feather impressions (Charig et al.. 1986). A further point worth raising here is that any organic bonding material available to a forger in the 19th century for mixing cement would have shown some evidence of cracking or shrinking away. No such cracking or shrinkage has been observed.
ii) The presence of detailed feather impressions on the main slab, coupled with their absence on the general surface of the counterslab was taken as evidence that the cement layer on the counterslab was removed either because it was too difficult to match the feather impressions or that material fell off when the counterslab was hammered (Hoyle et al.. 1985; Watkins et al.. 1985b).
Fossils from the Solnhofen Limestone are commonly found concentrated on one part of the slab, with only a faint impression on the counterpart. This is because the part containing the bulk of the fossil represents the covering sediment, into which the body of the animal protruded. The faint impression on the counterpart represents the impression made on the lagoon bed where the animal came to rest on the bottom (e.g. Swinburne 1988). In other words, the bulk of the specimen lay project ing above the sea floor and was eventually encapsulated within the overlying bed. Upon splitting along the original sea floor surface, the bulk of the fossil will be preserved in the bed above the spilt, while the original bedding surface will retain only an impression. Thus the part containing the bulk of the fossil represents the overlying bed. This difference between a well preserved part and a poorly preserved counterpart is well known in Solnhofen fossils. Thus, using this differential preservation as a criterion for forgery means that all Solnhofen fossils must be suspect!
iii) The occurrence on the slabs of smooth, flattened, slightly elevated areas resembling "blobs of chewing gum" (Watkins et al.. 1985a, p. 265), only a few millimetres in length and not always matched by corresponding depressions on the opposite slab (Watkins et al. 1985b), some bearing faint but detailed feather impressions. This was claimed to be fragments of the lost cement layer which was not fully removed (Watkins et al.. 1985b). Hoyle et al.. (1985, p. 694) stated that these "blobs" are
"without any place to go should the main slab and counterslab be closed like the leaves of a book."
In other words they claimed that the slab would not fit tightly with the counterslab since the "blobs" did not correspond to any depression in the counterslab.
These "blobs of chewing gum" appear to be natural irregularities in the surface of the slab. Indeed,
"careful casting of the surfaces of both main slab and counterslab shows that there is always a good fit between the two, except where it has been destroyed by subsequent preparation. In no case is there an elevation on one slab 'without any place to go should the main slab and counterslab be closed like the leaves of a book.'" (Charig et al.. 1986, p. 623).
iv) The regularity of the side veins of the feather impressions was claimed to indicate a forgery, since the limestone could not have split so evenly as to break along the length of the feathers (Watkins et al.. 1985b).
The Solnhofen Limestone is well known for its smooth, level bedding planes along which it readily splits, providing an ideal, flat, smooth surface for use in printing and for exposing fossils. The extremely fine texture, essential for printing, is ideal in preserving the most delicate anatomical structures, such as the medusae of jellyfish, the hairlike setae of crustaceans and the wing membranes of pterosaurs (Charig et al. 1986; Barthel et al.. 1990).
v) The apparent "double strike phenomenon" was claimed to indicate that the same feather was printed twice in a slightly displaced position and was thus indicative of a forgery (Watkins et al. 1985a).
The reproduction of a double impression would be harder to forge that a simple single impression, thus making it unlikely that a forger would attempt such a double impression. Besides, it is also observed on the Berlin specimen and has recently been much more convincingly explained as representing two overlapping feathers (Rietschel 1985).
vi) The tail is in fact one large tail feather and the caudal vertebrae are in fact the central axis of the feather (Watkins 1985a).
Not only is the tail "obviously segmented" (Charig et al. 1986, p. 625), individual feathers can be seen attached to the vertebrae via ligaments (de Beer 1954; Charig et al. 1986).
Apart from the above comments, Charig et al. (1986, p. 623-624) cite further evidence against a possible forgery.
"Our conclusive evidence of the authenticity of the Archaeopteryx holotype, however, is provided by what appear to be a number of fine lines on the main slab that run in various directions across the feather impressions in the region of the forelimb; some of them extend through the bony elements ofthe skeleton and on to the tail. They are difficult to spot with the naked eye, but their presence is shown with great clarity by critically lit ultraviolet photography. Associated in a few places with the more easily visible linear staining of an orange-brown colour, they are presumably hairline cracks and are generally filled with mineral matter. These cracks are also present on the counterslab in precisely the same positions."
This indicates that there is no intervening cement layer between the two slabs, as does the presence of manganese dioxide dendrites which have grown over the feather impression in some areas. These too match precisely on the two slabs, even in microscopic detail (Charig et al. 1986).
Watkins et al. offered two reasons for the forgery, both implicating the then Superintendent of the Natural History departments of the British Museum, Richard Owen (Runyard 1985). Firstly they suggested that Owen forged the impressions to provide evidence in support of Darwin's ideas on evolution. Given Owen's hostility towards Darwin and his ideas (not towards evolution, merely towards Darwin's ideas on evolution) this is extremely unlikely. The other reason was that Owen laid a trap for Darwin, to tempt Darwin into making a fool of himself by declaring the fossil proof of evolution and then revealing the "evidence" to be a forgery. This however is ludicrous. Owen himself made a detailed description of the specimen (Owen 1863) thus laying himself and his reputation open to ridicule should the specimen prove to be a forgery. Besides, although Darwin knew of the specimen, he made only passing reference to it, describing it as " that strange bird Archaeopteryx" (Darwin 1866). He knew that one specimen could not prove his ideas on evolution.
In 1988 Spetner, Hoyle, Wickramasinghe and Magarits published a further piece concerning the analysis of two small rock samples taken from the London specimen, one from the feather region (called FM) and one from one side of the slab, away from the fossil (called MM). After the analyses were complete, several claims were made.
"Our contention is that the feather impressions were forged onto a fossil of a flying reptile." (Spetner et al. 1988, p. 15).
The only flying reptiles known (excluding Archaeopteryx and related forms) are the pterosaurs. However, Archaeopteryx does not possess the skeletal morphology to enable it to fly in the same way that pterosaurs did. Pterosaurs have been found in the Solnhofen limestone, complete with impressions of the wing membranes. No fossil reptile with a skeletal morphology similar to Archaeopteryx has been found with these membranes. Spetner et al. give no comment as to how this "flying reptile" managed to fly without the aid of feathers or, apparently, a membrane similar to pterosaurs. Indeed, it is likely that such a reptile could only have flown if it possessed feathers.
"The hairline cracks that they report to appear across the feathered region could have occurred naturally on the forgery premise. As the stone underwent slight movement, a crack in the underlying stone would tend to come through a thin layer of cement that had been placed on the surface." (Spetner et al. 1988, p. 16).
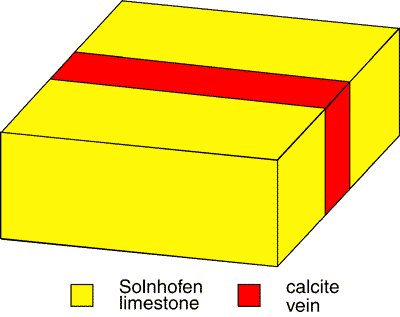
This fails to explain what is observed. The "crack" had been naturally infilled with calcite prior to the alleged forgery, thus the crack as a space to initiate upward propagation, no longer existed (fig 1).
Not only that but if the crack did propagate upwards, there would be a gap - within the crack - between the top of the calcite infilling the original crack and the top of the added layer of cement (fig 2).
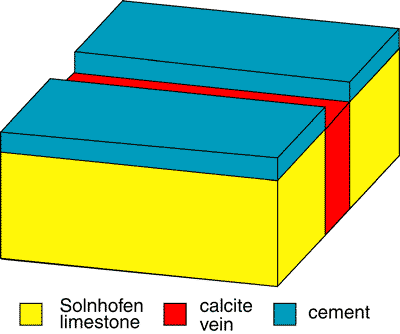
However, analysis of the slab shows the situation to be like that in Fig 1. So, in order for Spetner et al. to be correct, not only does the crack have to propagate upwards, but the infill has to mobilize and fill the gap between the old top of the slab and the top of the cement layer. This cannot happen merely by a "slight movement" of the slab. The presence of a number of these cracks in the configuration shown in Fig. 1, refutes the presence of a cement layer. The explanation in Spetner et al. (1988) is not viable.
"With regard to the claim that the dendritic patters appears on the feathered region, we can only say that they must have been mistaken. Fig. 1 shows the same dendritic pattern to which they referred, but it shows it in much greater detail than their figure. Fig. 1a is a photograph of the same area as shown by Charig et al. Fig 1b is a map of the area shown in Fig 1a to enable the reader to locate the detail in the photograph, the viewing of which may be facilitated by the use of a magnifying glass. In Fig 1, the dendritic pattern can be seen to lie wholly off the wing region, only abutting it on the patterns lower edge. The outline of the wing is indicated in Fig 1b by a heavy double line. Note that the dendritic pattern lies totally above the wing and just abuts it on the lower edge. . . . . The museum refuses us any additional access to the fossil so we have to make do with photographs that we already had. In any case, what is clear is that the dendritic patterns do not lie on the feathers as claimed by Charig et al." (Spetner et al. 1988, p. 16, original emphasis).
The claim that the dendritic pattern does not lie on the feathers is false. Fig 3b from Charig et al. (1986) clearly shows the dendritic pattern overlapping the feather impressions (note: this is clearest on the counterpart slab, fig 3b, not the slab figured by Spetner et al. - possibly because they did not photograph the counterpart slab?). Also Spetner et al.'s claim relied on their poor quality photographs of one area and a rather crudely drawn "map". Charig et al. (1986, p. 624) clearly stated that the dendrites "have grown over the feathers in places" (emphasis added). There is more than one occurrence. This, combined with the higher quality published photographs in Charig et al. (1986, fig. 3) clearly showing the dendrites overlapping the feathers, means that this refutation by Spetner et al. is unconvincing.
"Figs 4 are SEM photographs of the samples. A typical photograph of the MM shown in Fig 4a, which shows calcite grains varying in size from about 1 to 5 micrometres. These grains are typical of the fine carbonate structure one would expect to find in Solnhofen limestone. The FM, on the other hand can be seen to consist of a similar material with the addition of an unknown non-crystalline substance." (Spetner et al. 1988, p. 16-17)
It should be pointed out that their fig. 4 comprises 6 SEM photographs, one of the matrix material and 5 of the feather material. It is difficult to accurately compare the two samples since the SEM of the matrix material is shown at a higher magnification that those of the feather area (with the possible exception of figs. 4c and 4f which show close-ups of the "anomalous material", but these two have no scales included). Thus any apparent visual difference in the bulk make-up of the two samples (excluding the "anomalous substance") is an artifact of differential resolution. There appears to be no evidence of a fine binder of any kind in the rock matrix of the FM (feather) sample. In fact the matrix looks identical to the MM (matrix) sample.
"The unknown substance comes in various shapes. In some cases it has a rod-like appearance to which carbonate grains have adhered, as seen in Figs 4b and 4c. In other cases this substance has an odd shape as shown in varying magnifications in Figs 4d, 4e and 4f." (Spetner et al. 1988, p. 17)
The "rod-like" form is long and thin and hair-like. The "odd shape" is an amorphous, round blob, 0.0025 mm in diameter. The carbonate grains adhering to the unknown substance are extremely small, at 0.0006 mm.
"Note that the carbonate grains lie both above and below the non-crystalline substance. If the material were simply applied to the surface of the fossil as a preservative, as the museum proposes, one would expect to find it lying wholly above the carbonate grains." (Spetner et al. 1988, p. 17, original emphasis).
This claim is not supported by the photomicrographs. These show that the unknown material is clearly separate from the matrix carbonate. They appear to lie wholly above the carbonate grain matrix. Spetner et al. suggest that because the material is not at the surface, it cannot have been recently introduced except as part of a cement. However, the unknown material is clearly not forming part of any cement. Also the size of the sample (a few micrograms) means that the surface under examination is only a few 10ths of a millimetre below the original surface - although it is difficult to judge where the original surface was, since there is no indication of orientation in the photomicrographs - thus introduction into minute cavities in the surface cannot be ruled out. The carbonate grains which are coating the unknown substance are very small and were most probably dislodged during the cleaning and/or casting process, and adhered to the unknown material in the same manner that dust adheres to surfaces.
"An X-ray chemical analysis of the unknown substance in Fig. 4f is shown in Fig. 5. At low irradiation energy (10 kV) the analyser shows peaks of calcium, silicon, sulphur and chlorine. At higher irradiation energy (25 kV) peaks of zinc and lead appear. The intensity of the X-rays emitted from the elements detected in the unknown substance in the FM sample was relatively low. This low intensity suggests that the main chemical component of the unknown substance has an atomic number too low to appear in the analyses. Since our instrument does not detect elements whose atomic numbers are lower than that of sodium. We can only suspect that the unknown substance is principally carbon, and that it is likely to be of organic origin. The size of the sample that we received from the Museum did not permit a true chemical analysis in which organic substances could be identified." (Spetner et al. 1988, p. 17).
Their Fig. 5 is labeled, "X-ray luminescence results from amorphous body shown in Fig. 4f." However this is incorrect. The analysis was obtained using Energy Dispersive Spectrometry - "X-ray luminescence" is not a term that is used to describe X-ray analyses. The analysis produced peaks for calcium, as expected, but also for silicon, sulphur and chlorine (see Fig. 3) - not elements usually associated with organic matter.
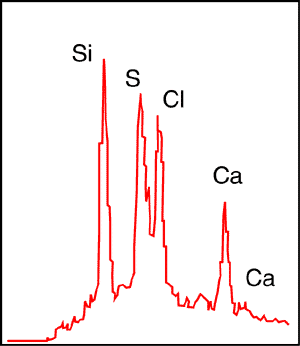
The sulphur peak must be regarded as suspect since the low resolution (140-150 ev) of energy dispersive spectronomy will fail to differentiate between a Ka sulphur peak and a Ka lead peak, especially at 10 kV. At 25 kV a lead peak appears, indicating the presence of lead and thus at least a portion of the "sulphur" peak, at 10 kV, must be regarded as lead (again a substance not associated with organic matter).
Several things should be pointed out at this point:
When these facts are taken into consideration, a much more likely explanation presents itself.
The "unknown substance" analysed by Spetner et al. was, most probably a fragment of silicone rubber which had becomed entrained just below the surface of the slab during its fluid phase. But would a piece of silicone rubber produce the X-ray spectrograph shown? Well, the chemical formula of silicon rubber is show in Fig. 4.
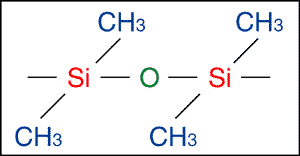
The large amount of silicon found in silicone rubber can explain the silicon peak seen in the spectrograph. As for the claim that the low intensity of X-rays emitted
"suggests that the main chemical component of the unknown substance has an atomic number too low to appear in the analyses. Since our instrument does not detect elements whose atomic numbers are lower than that of sodium. We can only suspect that the unknown substance is principally carbon, and that it is likely to be of organic origin." (Spetner et al. 1988, p. 17)
The presence of carbon, hydrogen and oxygen in silicon rubber (see Fig. 4), all with atomic numbers below that of sodium, can account for this, without the need to invoke organic matter.
But what of the chlorine peak? Well, the presence of chlorine is a problem if the material was of organic origin, but is no problem if the material is silicone rubber. Silicone rubber is commonly fixed with substances called Trichlorosilanes (SiCl3H) [no prizes for guessing what the "-chloro-" stands for! :-)], the mixing of which results in the locking together of long polymers strands of the rubber (Fig. 5).
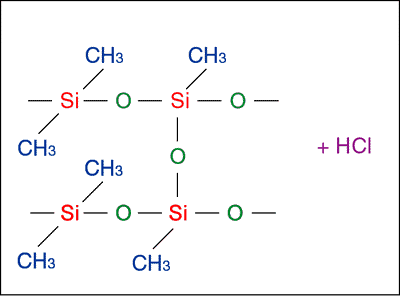
A byproduct of this reaction is HCl. Thus the chlorine peak could represent unreacted Trichlorosilane fixer, or a residue of HCl.
No evidence for the presence of an organically bound cement is presented. Photomicrographs indicate that there is no difference in the matrix from the fossil area as compared with the limestone. No organic matter was detected in the samples analysed. The presence of organic carbon is a suggestion presented by Spetner et al., but is one which is inconsistent with the chemical analyses presented in the article. These analyses suggest that anomalous material analysed by Spetner et al. is more likely to be silicone rubber from a mould prepared before the rock sample was taken, than an organic residue from - or included in - some sort of cement.
Hoyle and Wickramasinghe appear to still be confused over the preservational style of Archaeopteryx.
"The original piece of rock containing the genuine Compsognathus fossil was probably split with a bias of bones towards the main slab, but the bias could not have been so well marked as we see . . " (Hoyle & Wickramasinghe 1986, p. 51).
As Swinburne (1988) points out, the splitting is a fundamental property of the rock (it splits along a bedding plane, leaving the fossil mostly on one side. Also,
"it would be impossible we think for a bird-like creature to fossilise in such a way that the impressions of tens of thousands of feather barbs were all confined in an ordered way to a single surface, even if it be supposed possible to find that initially unknown unique surface working inwards from a boundary of a chunk of rock" (Hoyle & Wickramasinghe 1986, p. 59).
As explained by Swinburne,
"Hoyle and colleagues have misapplied the depositional theory, unaware of the attitude of the fossil. Archaeopteryx came to lie belly-up with wings outstreached on an extremely flat, well consolidated surface. It was covered by a pulse of suspended sediment which rained down into the stagnant pond in which it lay. The wings were then flattened against this plane by the weight of overlying sediment." (Swinburne 1988, p. 276).
This surface would eventually be the one which splits resulting in the assymetric preservation of the part and counterpart.
The evidence claimed by Watkins et al. to indicate that the feather impressions are a forgery appear to be easily explainable by natural processes. Detailed study of the London specimen both across the surface and in vertical section have failed to provide any evidence to support the contention that a layer of cement is present. The method claimed to have been used to produce the forgery cannot explain the presence of fine lines crisscrossing the fossil, or the matching dendrites on the slab and counterslab, which occur on top of the feather imprints. The feather imprints on the Maxberg specimen, despite claims to the contrary, are clearly identifiable as such. In this case, forgery of the type envisaged by Watkins et al. can be discounted because of the fact that the impressions run underneath the bony elements of the skeleton.
Something that should be obvious to anyone is that
"any conclusions about the authenticity of the fossil should be based on the best possible evidence. Photographs are just one ingredient of such evidence" (Parmeter & Greenaway 1985, p. 458).
Watkins et al., however, cite as evidence of their claims a set of "rudimentary," "poor" photographs having "too much contrast and too soft a focus," without looking at the much more extensive and better quality Museum photographs.
The claims that the feathers of Archaeopteryx are fake has been shown to be unsupported. Thus the claim that "the significance of Archaeopteryx lies in the fact that it represents the only unquestionable case of a fossil showing a transition between two vertebrate classes, aves (birds) and reptilia (reptiles)" has been upheld. In other words, Watkins et al. claim that Archaeopteryx represents a transitional form, but cannot be accepted as such because it is a forgery. Since the claim of forgery has not been substantiated, Archaeopteryx must therefore be an example of a transitional form by Watkins, et al.'s own admission (notwithstanding the fact that they mischaracterise Archaeopteryx as the "only" case).
I doubt however, that this particular quote will show up in any creationist literature.
Thanks go to Rich Trott for suggesting improvements. The text was originally coverted to HTML format for the talk.origins archive by Brett Vickers.
This is a University of Ediacara Palaeontological Contribution.
Barthel, K.W.; Swinburne, N.H.M. & Conway Morris, S. (1990) Solnhofen: A Study in Mesozoic Palaeontology. Cambridge University Press, Cambridge. 236pp. ISBN 052133344X.
de Beer, G. (1954) Archaeopteryx lithographica A study based on the British Museum specimen. British Museum, London. 68 pp.
Benton, M.J. (1990) Vertebrate Palaeontology: biology and evolution. Unwin Hyman, London. pp 377. ISBN 0045660018
Charig, A.J.; Greenaway,; F. Milner, A.N.; Walker, C.A. & Whybrow, P.J. (1986) Archaeopteryx is not a forgery. Science, 232: 622-626.
Colbert, E.H. & Marales, E. (1991) Evolution of the vertebrates: a history of the backboned animals. Wiley-Liss, New York. pp 470. ISBN 0471850748
Crawley, G. (1985) Archaeopteryx photographic techniques (reply). British Journal of Photography, 132: 458.
von Heller, F. (1959) Ein dritten Archaeopteryx-Fund aus den Solnhofener Plattenkalken von hangenalttheim/Mfr. Erlanger Geologische, 31: 1-25.
Hoyle, F. & Wickramasinghe, N.C. (1986) Archaeopteryx The Primordial Bird. Christopher Davies Ltd., Swansea.
Hoyle, F.; Wickramasinghe, N.C. & Watkins, R.S. (1985) Archaeopteryx. British Journal of Photography, 132: 693-694.
Kemp, T.S. (1982) Mammal-like reptiles and the origin of mammals. Academic Press, New York. pp 363. ISBN 0124041205.
Ostrom, J.H. (1976) Archaeopteryx and the origin of birds. Biological Journal of the Linnean Society, 8: 91-182.
Ostrom, J.H. (1972) Description of the Archaeopteryx specimen in the Teyler Museum, Haarlen. Proceedings Koninklijke Nederlandse Akademie Van Wetenschappen, B, 75: 289-305.
Owen, R. (1863) On the Archaeopteryx of von Meyer, with a description of the fossil remains of a long-tailed species from the lithographic stone of Solenhofen [sic]. Philosophocal Transactions of the Royal Society of London, 153: 33-47.
Parmenter, T. & Greenaway, F. (1985) Archaeopteryx photographic techniques. British Journal of Photography, 132: 458.
Rietschel, S. (1985) Frankfurter Allgemeine Zeitung. 8th May 1985: 31.
Runyard, S.A. (1985) Minutes of meeting - 23rd May 1985.
Spetner, L.M.; Hoyle, F.; Wickramasinghe, N.C. & Magaritz, M. (1988) Archaeopteryx - more evidence for a forgery. The British Journal of Photography, 135: 14-17.
Swinburne, N.H.M. (1988) The Solnhofen Limestone and the preservation of _Archaeopteryx_. Trends in Ecology and Evolution, 3(10): 274-277.
Trop, M. (1983) Is Archaeopteryx a fake? Creation Research Society Quarterly, September 1983: 121-122.
Watkins, R.S.; Hoyle, F.; Wickrmasinghe, N.C.; Watkins, J.; Rabilizirov, R. & Spetner, L.M. (1985a) Archaeopteryx - a photographic study. British Journal of Photography, 132: 264-266.
Watkins, R.S.; Hoyle, F.; Wickrmasinghe, N.C.; Watkins, J.; Rabilizirov, R. & Spetner, L.M. (1985b) Archaeopteryx - a further comment. British Journal of Photography, 132: 358-359, 367
Watkins, R.S.; Hoyle, F.; Wickrmasinghe, N.C.; Watkins, J.; Rabilizirov, R. & Spetner, L.M. (1985c) Archaeopteryx - more evidence. British Journal of Photography, 132: 468-470.
Wellnhofer, P. (1974) Das funfte skelettexemplar von Archaeopteryx. Palaeontographica A, 147: 169-216.
Wellnhofer, P. (1988) Ein neuer Exemplar von Archaeopteryx. Archaeopteryx, 6: 1-30.
Wellnhofer, P. (1993) The seventh specimen of Archaeopteryx from the Solnhofen Limestone. Archaeopteryx, 11: 1-47.

Home Page |
Browse |
Search |
Feedback |
Links
The FAQ |
Must-Read Files |
Index |
Creationism |
Evolution |
Age of the Earth |
Flood Geology |
Catastrophism |
Debates