Botanik online 1996-2004. Die Seiten werden nicht mehr bearbeitet, sie bleiben als historisches Dokument der botanischen Wissenschaft online erhalten!
Interactions between Plants and Parasitic Fungi
The evolution of fungi is mainly dependent on the further development and spread
of green plants. The majority of fungal species is saprophytic, some few are parasitic.
The latter are at least in certain stages of their life cycle dependent on a particular
supply of nutrients (vitamins, for example) that can only be provided by living cells.
It is quite striking that fungal diseases are rather rare in nature due to the very
effective defence mechanisms of plants that inhibit a spread of parasitic fungi.
The solid cell wall including all its introsusceptions and appositions (the cuticle,
for example) that inhibits the penetration of the tissues and cell lumina by fungi,
bacteria, viruses, etc. is the first line of defence. Additionally, a wide spectrum
of secondary defence compounds exists, many of which are fungicides and/ or bactericides.
These substances are often not produced until infection occurs (i.e. until their
production is induced, example: phytoalexines).
It is remarkable that plant parasites (and this is equally true for fungi,
bacteria, and viruses) are strictly host-specific.
Some of them depend on an alternation of hosts, so that one stage of the parasite’s
development occurs in one host, the following in another that is phylogenetically not
closely related to the first host. Parasites cause far larger damages in monocultures
(agriculture) than in plant communities rich in species due to their high host
specificity. The annual loss of foodstuff caused by them corresponds to an amount
large enough to provide 300 million people.
Parasitic fungi have at least three strategies to get hold of plant ingredients:
They produce enzymes for the breakdown of cell walls and cuticles.
They produce toxins that do either reduce the activity of the host cells or inhibit it completely.
They produce plant-specific substances (hormones, for example) thus meshing
with the hormonal equilibrium of the plant cell and causing disruptions of the
growth and differentiation of the cells and tissues. Gibberella fujikuroi,
for example, secretes gibberellins that influence the growth of the host plant (rice).
Studies of this fungus lead to the detection of the gibberellins (class of phytohormones).
The literature about parasitic fungi is very extensive. Many studies originate
in economic considerations. The majority deals with the classification of the fungi,
their life cycles, the symptoms of the plants and their diagnostic, the host range,
and the search for the host’s factors of resistance. The plants’ molecular mechanism
of action after infection with fungi has by now just been elucidated in a few cases,
and these examples can only partially be generalized due to the variety of possible reactions.
The resistance of the hosts is not only based on the general and unspecific
defence mechanisms mentioned above, but also on the production of specific,
genetically determined substances directed against specific fungi. Genetic
analyses showed that the resistance is on one hand caused by the existence
of dominant alleles of the respective genes, and on the other hand by resistance
genes that are inherited independently of each other.
Some Remarks about the Classification of Parasitic Fungi
It is distinguished between two types of parasitism:
 |
Necrotrophic Parasitism:
the infection leads to a destroying of the tissue and thus to the death of the plant.
The fungi are often only optionally parasitic, they can just as well proliferate in dead
or withering plant material.
|
 |
Biotrophic Parasitism: In this case,
parasite and host live together for longer periods of time. The parasite takes
nutrients and growth-regulating substances from the plant, but does not kill it.
Most biotrophic fungi are obligatory parasites. They survive only limited
saprophytic phases; especially the development of a fruiting body is dependent
on the presence of a host. The cultivation of single (vegetative) stages of these
fungi in cell-free nutrient medium succeeded only in a few exceptions.
|
Parasitic fungi can be found in all classes of fungi, the hosts in all systematic
groups of plants (and blue-green algae). The kingdom of fungi subdivides into
myxomycetes or moulds (slime fungi) and
eumycetes, the higher fungi.
One of the economically most important representatives of the parasitic myxomycetes
is Plasmodium brassicae, the causative agent of clubroot, a disease whose symptoms
can be seen in the roots of numerous species of cruciferes. The changed differentiation
pattern of the infected host tissue is caused by a 50- 100 fold increased auxine content,
a 10- 100 fold rise of the cytokinin content, and an increased degree of ploidy of the
respective tissues’ nuclei (due to the high level of cytokinin maybe?; I. C. TOMMERUP and
D. S. INGRAM, 1971).
Spongospora subterraneae is the causative agent of the potato’s
powdery scab that can serve as a vector for the potato-mop-top-virus.
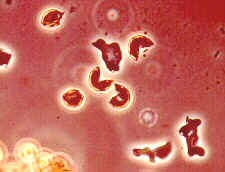


Myxostelida - resting spores splitting open to release myxamoebae - Trichiales - young sporangia of Trichia decipiens - Trichiales - plasmodiocarp of Hemitrichia serpula
© Bryce KENDRICK
Eumycetes
.
Phytophthora-species need among a few others Solanaceae (species of the potato family) as hosts.
Phytophthora infestans is the causative agent of late blight of potato.
The early destruction of the foliage and the reduction of the rate of photosynthesis
caused by it leads to heavy crop failure. The host spectrum is - in contrast to some
other Phytophthora-species (potato, tomato, and a few others) - very restricted.
The fungus was the cause of a big famine in Ireland in the middle of the
19th century (1845/ 47). A huge migration into the USA was the consequence.

Four kinds of Ascus exist. They do not look alike.
They come in two basic kinds,
called unitunicate and bitunicate.
The picture illustrates the bitunicate type.
© Bryce KENDRICK
|
Ascomycetes
. Numerous species like
Taphrina insititae (host: plums and others), T. betula (birch),
T. cerasi (cherry), T. deformans (peach) and others that cause
leaf curls in different hosts belong to the ascomycetes. Most of these species
develop haustoria (exception: T. deformans), i.e. processes into the inner
of the host cell lumen.
The most important precondition for the development of a haustorium is a local
perforation of the plant cell wall. The growing haustorium widens into a vesicle-like
structure after having penetrated the cell lumen. The host cell’s plasmalemma is not
penetrated. Haustoria do therefore not grow into the plasma of the host cells.
Electron microscopic studies of different types of haustoria showed that the
plasmalemma changed structurally, though. It folds up and a lot of electron-dense
material is incorporated into the membrane. These observations point to an active
defence reaction of the cells. Leaf curls are caused by an increased, but uneven
growth of single leaf areas. Experimental data suggest that the leaves’ content
of cytokinins and IAA is increased.
The species causing witches’ broom in numerous host trees belongs to the Taphrina
family, too. The symptom is produced by an increased number of vegetative points that
are the reason for an irregular, tuft-shaped branching pattern of the infected plant parts.
 The order Erysiphales consists of the family Erysiphaceae
(mildew).
Mildew is the collective name for a variety
of Erysiphacae-species that live parasitically on a number of angiosperms:
Erysiphe graminis (corn mildew),
E. communis (on pumpkin), and E. polygoni (on peas, clover,
and other Leguminosae). The fungal mycelium spreads usually on the upper or
the undersurface of the leaf. A few epidermal cells are penetrated by occasional haustoria.
The order Erysiphales consists of the family Erysiphaceae
(mildew).
Mildew is the collective name for a variety
of Erysiphacae-species that live parasitically on a number of angiosperms:
Erysiphe graminis (corn mildew),
E. communis (on pumpkin), and E. polygoni (on peas, clover,
and other Leguminosae). The fungal mycelium spreads usually on the upper or
the undersurface of the leaf. A few epidermal cells are penetrated by occasional haustoria.
Numerous subspecies of Erysiphe graminis have specialized on cereals.
Some species, for example, grow on wheat but not on barley, while the opposite is
true for others.
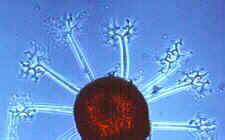
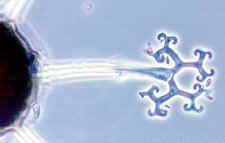
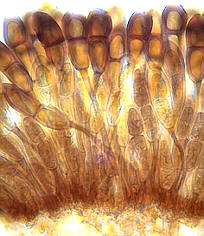
Erysiphales - SEM of conidial chains and mother cells of the Oidium anamorph of Erysiphe on a host leaf (picture to the left) -
dichotomously branched ascomatal appendage of Microsphaera - ascoma of Microsphaera, with dichotomously branched appendages
© Bryce KENDRICK
Ceratocystis ulmi and related species cause the Dutch elm disease.
The disease was discovered in Britain in 1927, where 10% of all elms were
infected between 1930 and 1940. In the dry summer of 1947, it spread epidemically
in Germany, too.
Botrytis cinerea (grey mould) is a not much specialized parasite.
It infects lettuce leaves as well as a variety of juicy fruits (tomatoes,
strawberries, etc.) if the weather is humid enough.
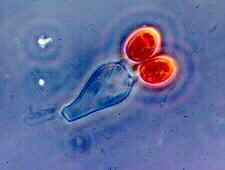
Basidium of a sequestrate basidiomycete, Gastrocybe, in which the basidiospores are symmetrically mounted, and are not forcibly discharged
© Bryce KENDRICK
|
Basidiomycetes. Two important orders of parasitic fungi
belong to the basidiomycetes: Ustilaginales (smuts) and Uredinales (rusts). The order
Agaricales consists mainly of the already discussed mycorrhizal fungi, only few species
are parasitic (honey fungus, for example). More than 1000 species of Ustilaginales live
parasitically on hosts of more than 75 angiosperm families. They produce dark, powder-like
traces consisting of fruiting bodies strung together on leaves, shoots, flowers, and fruits.
They develop usually no haustoria. Their extensive mycelium spreads through the
intercellular spaces of the host plants. Uredinales include about 4,000 species in 100 genera.
They are characterized by the red-brownish colour (rusts !) of their spores.
Smuts develop haustoria and infect a range of angiosperms, gymnosperms, and pteridophytes.
Puccinia graminis is the classic example of a fungus with an alteration of hosts.
Its haploid (monocariontic) mycelium grows on Berberis (barberry), its dicariontic
mycelium on different grasses. P. graminis is characterized by a complex alternation
of generations. Up to five different types of spores can be generated during its course.
P. graminis can live on grasses for an infinite time in its dicariontic state,
if barberry is not present.

Uredinales. Teleutosorus. The stalked,
two-celled winter spores (teleutospores) develop on cereals after the change of the host
in summer and hibernate on the ground. These probasidia develop into the original,
septate basidiospores (photo: W. KASPRIK).
The preconditions for this change, however, are mild winters, since the dicariontic, asexual
uredospores (summer spores) cannot survive long and cold winters. Wind can on the other
hand disperse fungus spores thousands of kilometres so that a local extermination of
barberry provides no lasting protection against Puccinia. This species consists
of a number of subspecies, too. One has specialized on wheat, another on oat, a third
on rye, etc. Puccinia graminis belongs to the few parasitic fungi that can be
cultivated cell-free on agar (under addition of yeast extract).

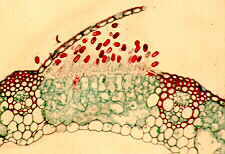

Uredinales - uredinia of Puccinia graminis tritici erupting through epidermis of wheat (Triticum). - - vertical section through a uredinial sorus of Puccinia graminis tritici in a wheat leaf. - vertical section through an aecium of Puccinia graminis tritici.
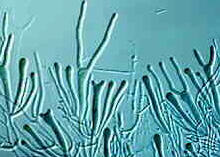
Dacrymycetales - "tuning fork" basidia of Dacrymyces at various stages of development
© Bryce KENDRICK
Fungi imperfecti. This is the collective name
for fungus species that do not develop fruiting bodies and that are therefore rather
difficult to classify. Because of ultrastructural studies, it is assumed today that
most of them belong to the ascomycetes. Several species have been used as test objects for
studying molecular processes occurring during the infection of plants. We will therefore
discuss them two subchapters later.
Parasitic aquatic fungi
are rather unknown to the public, since they are of no economic importance.
Only few scientists have studied them intensely.
More than 200 filamentous marine fungi have been described. Between a quarter
or a third of them live parasitically on algae. Most of them belong to the ascomycetes.
Brown, red, and green algae as well as diatoms serve as their hosts.
Lagenisma coscinodisca, a diatom parasite, has been especially well documented
with the help of the electron microscope.
Most parasites of freshwater algae belong to the chytrids that were
discovered by A. BRAUN (Berlin) in 1856. He described them as follows:
" The whole little plant consists of a simple, vesicle-like cell
that does often penetrate the cells of the nutritive organism with a
root-like excrescence."
Chytrids develop no hyphae, but a non-cellular
and partially rather extensive system of rhizoids that seizes the host cells and can
even enclose them. Nearly every known species of algae has its own specific parasite.
The fungus infections should not be underestimated. Infections of nearly all algal
cell-colonies have often been observed towards the end of a ‘water bloom’, i.e.
the massive growth of an algal species.
Chytrids infect both blue-green algae (Cyanophyceae) and Volvocales,
as well as Chlorococcales. These three groups of algae differ in the chemical composition
of their cell walls so that the respective parasites have to have the most diverse enzyme
systems at their disposal. The destruction of a cell occurs only after a cell-to-cell
contact has been established, i.e. the fungus secretes no lytic substance into the
adjacent medium that destroys all cells of a colony that are surrounded by
a gelatinous coat.
Years of research into the population dynamics of diatom and cyanophycean parasites showed,
that the fungus infection is able to reduce the number of individuals drastically, especially
at the beginning of a population’s growth, and to accelerate the decline of a population
lastingly. The studies took place in South England’s Lake District. A reduction in the number of
individuals of an algal species is usually compensated by an increase in the population
size of another species (H. M. CANTER, Freshwater Biological Association, Ambleside).
Such quantitative analyses and a larger attention to the consequences of the fungus
infection offer possible explanations for the fact that a species of algae is dominant
in one year and does scarcely, or not at all occur in the following year.
The infection of algal colonies with chytrids is not the only reason for the
disintegration of a water bloom. Other reasons, like the lack of nutrients
(especially of phosphate) or weather-caused changes are often decisive,
especially if the fungus infection is limited and only few cells of a colonyare destroyed
(as was shown e.g. for the end of a bloom of the blue-green alga Microcystis aeruginosa)
© Peter v. Sengbusch - Impressum