Botany online 1996-2004. No further update, only historical document of botanical science!
Research into photoperiodism began more than 60 years ago when the existence of short-day and long-day plants was discovered. Its analytical stage started after botanical institutes had been equipped with climatic chambers where plants could be cultivated under controlled light and temperature programs. It turned out that it is not the day length (light period) that is decisive for stimulation of flowering but the dark period. A minimal light period is required for the production of enough assimilates. Moreover can the dark period exert both a stimulating and an inhibiting influence on flower formation. Both effects can be reversed by permanent weak light.
In 1945 documented M. W. PARKER, S. B. HENDRICKS, H. A. BORTHWICK and N. J. SCULLY the action spectrum of the photoperiodic reaction. They did even then point out the importance of red light. Later on did it turn out that photoperiodism, too, is controlled by the phytochrome system.
K. C. HAMNER (University of California, Los Angeles (UCLA)) detected that the flower formation of the short-day plant Biloxi soja (soy bean) is dependent on the ratio of dark period to light period. After a light period of eight hours are dark periods of 24 hours or a multiple of 24 hours optimal. It is not necessary that a dark period follows a light phase immediately. A short interruption by interference light is enough. The short-day plant Kalanchoe blossfeldiana does not flower if the interference light is offered at the wrong time.
With the onset of a light period begins a physiological activity called the photophilic stage. After 9-12 hours of light is the development of the plant inhibited by all further exposure to light: the plant enters its skotophile, i.e. its darkness-loving phase. The next photophilic phase begins just a few hours later, long before dawn and thus quite independent of the actual conditions of alternation of day and night. A reduced light program where the plant receives just a few hours of light every 72 hours is enough to synchronize the alternation of skotophile and photophilic phase. It should be noticed that the endogenous rhythm contributes decisively to the maintenance of an approximate 24-hour rhythm also called the circadian rhythm. Photoperiodism controls mainly the relative share the two mentioned phases have during a 24-hour period.
A model outlines the processes that take part in a short-day plant during light and darkness. It shows that the stock of PFR is exhausted by an enhanced breakdown and the lacking supply at the end of a photophilic phase so that the plant is not able to perceive light any more.
Since growth and differentiation are dependent on light do their physiological activities stagnate. They need a dark period during which the pool of PR is replenished. This pool does not grown unlimited but is broken down again during long periods of darkness so that the skotophile phase returns periodically. The measurement of time is coupled to a circadian rhythm, not to the phytochrome system. Animals, too, have a photoperiodism but no phytochrome system. They measure time with other methods.
The differences between short-day and long-day plants in view of the effect of phytochrome can be summarized as follows (according to H. MOHR and P. SCHOPFER, 1978):
long-day plants
: short day + enough PFR (during the middle of the skotophile phase) > production of flowering hormones
short-day plants
: short day + enough PFR (during the middle of the skotophile phase) > no production of flowering hormones
The qualitative difference in the reaction to the same signal is genetically fixed.
No other botanical substance has so long and so unsuccessfully been searched for as for the flowering hormone or florigen. It did at least became a name, and its existence could be proven by grafting experiments. Phytochrome that is localized within the leaves is required for the control of flower formation. The conditioning, i.e. the signal necessary for stimulation of flowering or for suppression of flower formation is also generated within the leaves. The majority of the classic grafting experiments was performed with Nicotiana-species. Nicotiana sylvestris is a long-day plant and Nicotiana tabacum, var. Maryland Mammoth (M.M.) is a short-day plant. Most other Nicotiana tabacum-species are day-neutral.
If Nicotiana sylvestris, for example, is cultivated under short-day conditions, does it not develop flowers, but if a leaf of a Nicotiana tabacum, M.M. plant that was cultivated under short-day conditions is grafted to Nicotiana sylvestris, then it is stimulated to flower. This means that the Nicotiana tabacum, M.M.-leaf had produced a substance that was transferred to the recipient after grafting and that caused its flower formation. This proved the existence of a florigen. The opposite experiment works, too: a Nicotiana tabacum, M.M.-recipient flowers under long-day conditions under the influence of a Nicotiana sylvestris graft. Similar results are received when grafting Nicotiana sylvestris, M.M. and a species of the genus Hyoscyamus (a long-day plant). Experiments with other species from several genera yielded comparable results. All results can be combined in consistent scheme.
In summary can the following conclusions be drawn (A. LANG, MSU-DOE Plant Research Laboratory, East Lansing, 1984):
In the leaves of plants is a hormone-like substance (florigen) produced, maybe also a complex of substances, that is conducted to the shoot meristems in order to stimulate them to pass from vegetative growth to flower formation.
The florigen is not species-specific. It can be transferred to members of the same species, to members of one genus and to members of different genera. The transferability seems to be limited only by the acceptability of the graft. A good connection between the phloem of the two partners seems to be important. Grafting of monocots are very difficult to do and a florigen transfer by grafting has not been successful, but it could be shown with other techniques that it is required in monocots, too.
Florigen is physiologically not specific. It can without difficulty be exchanged between short-day, long-day and day-neutral plants. It is very likely that it is identical in all plants. The main difference between short-day and long-day plants is that the florigen production occurs only under a certain (inductive) light program, that differs in the two types.
It seems that a likewise transferable substance exists in several long-day plants that is produced under short-day conditions and suppresses flower formation. It is called antiflorigen. The reaction is weak and cannot be detected in all combinations. Florigen and antiflorigen appear to be antagonists. The flower formation is mainly based on the ratio of the two substances.
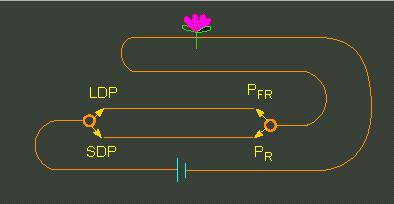
How can the combined action of florigen and phytochrome be explained? A. LANG illustrated it by the analogy of a circuit with alternative parallel connection. Accordingly has the ‘phytochrome switch’ of a long-day plant that is developing flowers to be on PFR, that of a short-day plant under the same condition on PR. In this simple way can the scheme explain the production or non-production of florigen. Both connections would have to be functional in day-neutral plants.
Contrary Reaction of Long-Day and Short-Day Plants to Daylength
The model is an analogy of a circuit with two wires and two switches. To the left: the genetically controlled switch, to the right the phytochrome switch. The latter has to be on PFR in a long-day plant and on PR in a short-day plant (A. LANG, 1984).
© Peter v. Sengbusch - Impressum