Botany online 1996-2004. No further update, only historical document of botanical science!
It has already been mentioned that a system of vascular bundles runs through all higher plants. It evolved as a response to the increase in the size of plants, which caused an progressing separation of roots and leaves in space. Thus, plants developed systems for long distance transport - that is, translocation. In that way became the exchange of the products of absorption and assimilation possible. The system consists of xylem and phloem. The xylem is the tissue that translocates water and minerals. Transport occurs acripetally, i.e. in the direction of the shoot's tip. The phloem is the tissue that translocates assimilates from mature leaves to growing or storage organs and roots. It serves, too, to redistribute water and various other substances It may run either basipetally, i.e. in the direction of the root or bidirectionally (both acripetally and basipetally).
The xylem of a living plant is an interconnected, water-containing apoplastic system of communicating tubes in which the water holds together by cohesion forces (J. BÖHM[ 1831 - 1893], Hochschule f. Bodenkultur, Wien). The xylem transports water and dissolved ions. The main portion of the water is taken up by young roots. Absorption occurs directly by the rhizodermis, and the area that takes up water and ions is enlarged by the formation of root hairs. From there flows the water through the cortex that may be developed as an exodermis, through the endodermis, the innermost layer of cortical cells and into the cells of the stele's vascular bundles. It is then transported through the roots to the shoot and finally into the leaves where it is given off by transpiration.
The way the water takes through the root tissue is decisively influenced by the structure of the involved tissues. Principally exist three ways to come through a tissue:
- apoplastically, i.e. through the cell walls,
- symplastically, i.e. from protoplast to protoplast via plasmodesmata
- through the cells (transcellular transport), i.e. from cell to cell during which the vacuoles are passed.
Outside the endodermis are all three ways equally favourable. As soon as it reaches the endodermis does the water have to pass through the protoplast since the apoplastic way is blocked by an encrustation of the wall called the Casparian strip. In some plants is the Casparian strip located in the exodermis so that the apoplastic barrier works sooner.
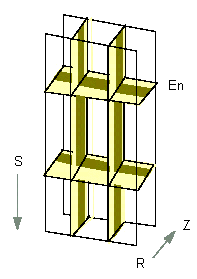
A Three-Dimensional Scheme of Root Endodermis Cells (longitudinal section). The Casparian strip is shown in dark olive. En: endodermis, S: direction towards the root tip, Z: stele, R: bark (according to U. LÜTTGE and N. HIGINBOTHAM, 1979).
The force that drives the water through the root is based on differences in the water potential of the root's surrounding (usually soil) and its xylem sap. The ions that have passed through the endodermis are usually trapped and cannot leave the stele any more. Due to osmosis builds thus a pressure up in the xylem called the endosmotic root pressure that presses the water with the dissolved ions upwards. Some plants have water stomata (also called hydathodes) or stomata at their leaf margins that secrete the water as droplets, a process called guttation. The root pressure works especially efficient at night but efficiency decreases during the day since the root pressure becomes much smaller than the rate of transpiration. Instead becomes the evaporation of water at the plant's surfaces important. The temperature difference between plant and the surrounding atmosphere causes water to vaporize. The resulting hypotension within the tissue is compensated for by water supplied by the root. Transport velocity depends on the water potential and the energy provided by the plant's surrounding, like solar radiation. The water potential itself is caused by the difference in atmospheric (low) and ground water content (high).
Besides the products of photosynthesis transports the phloem phytohormones, amino acids, herbicides, viruses and other substances. Assimilates are especially delivered to the storage organs of roots, fruits and seeds and to growing organs. Phytohormones travel both in the direction of the shoot's tip (acripetal direction) and towards the roots (basipetal direction) through the phloem and some viruses have been shown to use the phloem for their distribution throughout the whole plant. Phloem transport velocities of 30 - 150 cm/hour have been measured which means that a 0.5 cm long sieve element with an average flow of 90 cm/hour reloads every two seconds.
The use of radioactively labelled substances and autoradiographic methods allows the monitoring of solute translocation in plant tissues. While the mechanisms of xylem transport can rather simple be explained by warmth and water potential is the driving force behind phloem translocation less obvious. Phloem translocation has two aspects:
- phloem loading and
- transport itself.
The loading of phloem or, more precise, of the leaf sieve cells with assimilates is selective and requires ATP. It seems as if several different membrane-bound pumps perform this task. Sucrose accumulates faster than other sugars, and in some plant species are certain amino acids transported actively into the phloem. The active transport causes an osmotic pressure within the sieve tube elements that is about twice as high as that of the surrounding cells of the vascular bundle's sheath. Sugars are mainly responsible for this difference.
Model of Phloem Looading. The transport processes occur at the membrane of the sieve-elements (S = sucrose, C = carrier *, z = intermediates). The sugar transport occurs as a co-transport of sugar and potassium ions (according to R. T. GIAQUINTA, 1983)
The mechanism of phloem translocation was a subject of research from the 1930s to the mid-1970s. Today is the pressure-flow hypothesis first proposed by ERNST MÜNCH (Forstbotanisches Institut, Universität München) in 1926 widely accepted as the most probable mechanism. It states that the flow of solution in the sieve elements is driven by an osmotically generated pressure gradient between source and sink tissue. The gradient is a consequence of phloem loading at the source and phloem unloading at the sink. As has been mentioned causes the active loading at the source a high osmotic pressure in the sieve elements. This pressure leads to an influx of water that increases the turgor pressure. At the sink end of the translocation pathway produces phloem unloading a lower osmotic pressure within the sieve elements. The drop in the osmotic pressure that becomes lower than that of the xylem leads to an efflux of water and consequently to a decrease of the turgor pressure within the sieve elements of the sink. The model implies that some of the water circulates throughout the plant between xylem and phloem pathways. The sieve-element contents are physically pushed forward by bulk flow similar to water flowing through an open tap. Water movement through the phloem translocating pathways is driven by the pressure gradient that depends ultimately on the active short-distance transport mechanisms involved in phloem loading and unloading.
© Peter v. Sengbusch - Impressum
