
Botany online 1996-2004. No further update, only historical document of botanical science!
The development of stable supporting elements has been an important prerequisite for the evolution of large terrestrial organisms. Animals have endo- or exoskeletons that correspond in function to the woody stems or trunks of plants. The architectural design of the plant's body of vegetation is very complex. Thin petioles carry heavy and flat laminas, stems support leaves, flowers and fruits. All plant organs are exposed to mechanical strains. Organs above ground follow the wind's drift. Their high elasticity lets them either return to their original position, or it makes them swing around an imaginative axis. Trunks are stable enough to resist the wind's pulling. They withstand pressure and are inflexible, although their projecting treetops provide the wind with a large target. The wind makes the upper plant organs and the trunk act like a lever, a large part of the force is hence exerted onto the roots, that anchor the plant in the soil. Other functions of the root are water and nutriment uptake.
The strength of tissues protects also against enemies. The hard shell of many seeds prevents a chewing to pieces or puncturing by animals and avoids that parasites like fungi or bacteria force their way into them.
The preceding topic mentioned the high water-content of plant cells that lends a high tension to plant tissues and is caused by the turgor. It supplies plant tissues with a certain stability. Its actual importance is seen best in wilting leaves or flowers after their water supply has been stopped. Extensive specialized supporting tissues exist only in vascular plants. Despite the existence of huge marine brown algae (seaweeds, like Macrocystis, Laminaria), not a single terrestrial alga, whose thallus raises more than a few cell layers above ground, is known. Vascular plants have up to three types of supporting tissue:
- The collenchyma, a tissue of living cells,
- the sclerenchyma, a tissue of nearly always dead cells, and
- the vascular tissue consisting of both living and dead cells. It is responsible for the transport and dispersal of water, nutriments and assimilates.
All three types are reviewed below.
The larger a vessel plant is, the higher is its content of dead cells. Dead cells are exceptions among bryophytes, but very common in flowering plants. They are usually elongated (prosenchymatous) cells, in parallel to the axis of the respective organ and often combined in sheaves, the fibres. The botanist H. v. MOHL from Tübingen recognized already in the 30th of the 19th century that all these fibres spring from normal, living cells.
Supporting tissues reside generally in the periphery of plant organs. If the cells are combined in layers, tubes, whose stability is much greater than that of sticks of the same diameter are formed. The supporting tissues of ribbed or edged stems are concentrated in these ribs or edges. In submerse living vascular plants, the supporting tissue is reduced to a minimum.
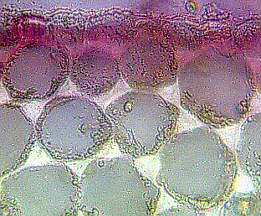
The collenchyma is the typical supporting tissue of the primary plant body and growing plant parts. Its prosenchymatous cells are living at maturity and are always kept in a primary state, which means that they are never lignified. Collenchyma walls are interspersed with groups of pits that tend to be organized in special areas. The name collenchyma derives from the Greek word "kolla", meaning "glue", which refers to the thick, glistening appearance of the walls in fresh tissues.
The collenchyma is the typical supporting tissue of the primary plant body and growing plant parts, though it is kept with unaltered structure and function even in outgrown organs like stems, petioles, laminae or roots. In cross-sections of stems, the collenchyma commonly appears as discrete strands or as a peripheral cylinder that lies, depending on the species, either directly beneath the epidermis or is separated from it by several layers of parenchyma. The cylinder is usually composed of several layers. Collenchyma is also found bordering the veins of dicot leaves. It forms fibres in edgy stems that run along the edges or ribs. Often either phloem or xylem of the vascular bundles is associated with collenchyma cells.
Many transitions prove the collenchyma's origin from the parenchyma. The differentiation is reversible, a degeneration to meristematic states has often been observed. The walls of collenchyma cells are strengthened by the deposit of cellulose and the coating with pectin. These strengthenings are often restricted to single parts or edges of the cell. The walls of parenchyma cells are opened by pits that are often arranged in special areas.
The unevenly thickened cell walls led the German botanist C. MÜLLER (1890) to distinguished between different collenchyma types:
- Angular Collenchyma. A thickening of the cell's edges can be seen in cross-section. Longitudinal sections show the elongated shape of both cell and thickening. A cross-section through the stem of Begonia rex or related species is the typical specimen used in botanical microscopic courses. Angular collenchyma occurs also in species of the following genera: Ficus, Vitis, Ampelopsis, Polygonium, Beta, Rumex, Boehmeria, Morus, Cannabis, Pelargonium and others.
- Tangential Collenchyma. The tangential walls of this collenchyma type are thicker than the radial walls. Examples: Sambucus nigra, species of the genera Sanguisorba, Rhoeo, Eupatoria.
- Lacunar Collenchyma. While hardly any intercellular spaces exist in the two types above, are those of this type very large. Clear gaps can be recognized between the cells. Occurrence: species of the genera Lactuca, Salvia, Prunella and the Composite-family.
The cell walls of collenchyma cells are distortable when stretched. Shape and arrangement of the cells cause a high mechanic stability with a capacity of 10-12 kg/mm2. This quality is especially advantageous in growing plant organs. It enables the collenchyma cells to stretch in synchrony with the other cells without spoiling the toughness of the tissue. The new state is stabilized by the simultaneous working-in of additional wall material.
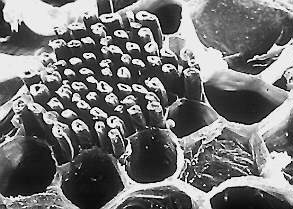
The other true supporting tissue is the sclerenchyma. Two groups of sclerenchyma cells exist: fibres and sclereids. Their walls consist of cellulose and/or lignin. Sclerenchyma cells are the principal supporting cells in plant parts that have ceased elongation. Sclerenchyma fibres are of great economical importance, since they constitute the source material for many fabrics (flax, hemp, jute, ramie).
Contrary to the collenchyma, mature sclerenchyma is composed of dead cells with extremely thick cell walls (secondary walls) that make up to 90% of the whole cell volume. The term "sclerenchyma" is derived from the Greek "scleros", meaning "hard". It is their hard, thick walls that make sclerenchyma cells important strengthening and supporting elements in plant parts that have ceased elongation. The difference between fibres and sclereids is not always clear. Transitions do exist, sometimes even within one and the same plant.
Fibres are generally long, slender, so-called prosenchymatous cells, usually occuring in strands or bundles. Such bundles or the totality of a stem's bundles are colloquially called fibres. Their high load-bearing capacity and the ease with which they can be processed has since antiquity made them the source material for a number of things, like ropes, fabrics or mattresses. The fibres of flax (Linum usitatissimum) have been known in Europe and Egypt since more than 3000 years, those of hemp (Canabis sativa) in China for just as long. These fibres, and those of jute (Corchorus capsularis) and ramie (Boehmeria nivea, a nettle), are extremely soft and elastic and are especially well suited for the processing to textiles. Their principal cell wall material is cellulose.
Contrasting are hard fibres that are mostly found in monocots. Typical examples are the fibres of many Gramineae, Agaves (sisal: Agave sisalana), lilies (Yucca or Phormium tenax), Musa textilis and others. Their cell walls harbour, besides cellulose, a high proportion of lignin. The load-bearing capacity of Phormium tenax is as high as 20-25 kg/mm2 and is thus the same as that of good steel wire (25 kg/ mm2). But the fibre tears as soon as it is put too great a strain on it, while the wire distorts and tears not before a strain of 80 kg/mm2. The thickening of a cell wall has been studied in Linum. Starting at the centre of the fibre are the thickening layers of the secondary wall deposited one after the other. Growth at both tips of the cell leads to simultaneous elongation. During development do the layers of secondary material seem like tubes, of which the outer one is always longer and older than the next. After completion of growth the missing parts are supplemented, so that the wall is evenly thickened up to the tips of the fibres.
Fibres stem usually from meristematic
tissues. Cambium and procambium are their main centers of
production. They are often associated with the xylem of the vascular
bundles. The fibres of the xylem are always
lignified. Reliable evidence for the fibre cells' evolutionary origin
of tracheids exists. During evolution the strength of the cell walls
was enhanced, the ability to conduct water was lost and the size of
the pits reduced. Fibres that do not belong to the xylem are bast
(outside the ring of cambium) and such fibres that are arranged in
characteristic patterns at different sites of the shoot.
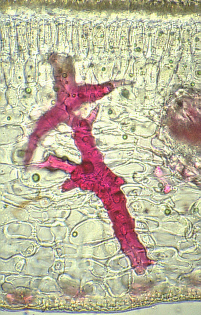
Sclereids are variable in shape.
The cells can be isodiametric, prosenchymatic, forked or
fantastically branched. They can be grouped into bundles, can form
complete tubes located at the periphery or can occur as single cells
or small groups of cells within parenchyma tissues. But compared with
most fibres sclereids are relatively short. Characteristic examples
are the stone cells (called stone
cells because of their hardness) of pears (Pyrus communis) and
quinces (Cydonia oblonga) and those of the shoot of the wax
plant (Hoya carnosa). The cell walls fill nearly all the
cell's volume. A layering of the walls and the existence of branched
pits is clearly visible. Branched pits such as these are called
ramiform pits. The shell of many seeds like
those of nuts as well as the stones of drupes like cherries or plums
are made up from sclereids.