
Botany online 1996-2004. No further update, only historical document of botanical science!
Assimilation tissues are - in a broader sense - all those tissues that are made from chloroplast-containing cells and are thus able to perform photosynthesis. They are found in all the green parts of a plant. An important aspect of photosynthesis is the integration of carbon dioxide into organic compounds. The resulting products are summed up as assimilates. The leaves of higher plants are by far the most important production centers - if you disregard unicellular aquatic algae. Leaves consist usually of three different tissues: the mesophyll, the epidermis and the vascular tissues. The mesophyll is a parenchyma tissue. It is a true assimilation tissue. In the leaves of most ferns and phanerogams, it is organized into palisade parenchyma and spongy parenchyma.
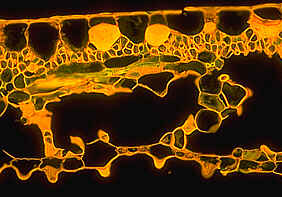
Cross-section through a Helleborus niger leaf. Semi-thin section of 3 µm thickness.
The cell walls are stained with calcofluorine
Before we advance to the details, it has to be understood that the terms mesophyll and assimilation tissue are not to be used as synonyms, since leaves devoid of chloroplasts exist that differ in organization only slightly from green leaves. They do thus have a mesophyll, but no assimilation takes place. Furthermore, assimilation may also occur, for example, in parts of the stem.
The "typical" leaf is of a
dorsiventral structure. The
palisade
parenchyma is usually directly
beneath the epidermis of the upper surface of the leaf. The
spongy parenchyma fills the space
beneath the palisade parenchyma. It is interspersed with a voluminous
intercellular system, whose cavities are in direct contact with the
atmosphere via the stomata. This roughly
outlined structure is always cited as being the prime example for
selection and adaptation. Our contemporary knowledge of
photosynthesis shows that an efficient assimilation tissue has to
meet the following criteria:
- It must make optimal advantage of the available light.
- The cells have continually to be provided with enough carbon dioxide (CO2).
- All cells must be connected via direct cell-to-cell contact with the vascular tissues in order to guarantee water supply and transport of assimilates.
 The
structure of the assimilation tissue of leaves fulfils these
requirements in a nearly ideal way. Additionally, the lamina is thin
and flat and the positioning of the leaves at the shoots is designed
to take maximal advantage of the light. The shadowing of chloroplasts
by other cells or leaves is thus reduced to a minimum. The supply of
the plant with light, carbon dioxide and, above all, water varies
strongly. It is therefore not astonishing that modifications
of this construction with enhanced adaptive values have developed
during evolution. Their organization is dependent on environmental
conditions.
The
structure of the assimilation tissue of leaves fulfils these
requirements in a nearly ideal way. Additionally, the lamina is thin
and flat and the positioning of the leaves at the shoots is designed
to take maximal advantage of the light. The shadowing of chloroplasts
by other cells or leaves is thus reduced to a minimum. The supply of
the plant with light, carbon dioxide and, above all, water varies
strongly. It is therefore not astonishing that modifications
of this construction with enhanced adaptive values have developed
during evolution. Their organization is dependent on environmental
conditions.
The cells of the palisade parenchyma are cylindrical. Neighboring cells look like the stakes of a palisade. The analogy is striking, when looking at the cross-section of a leaf, but is less suitable, when thinking of the three-dimensional arrangement of the cells. Cells of the palisade parenchyma contain three to five times as many chloroplasts as those of the spongy parenchyma. The chloroplasts stay usually near the cell's wall, since this adjustment guarantees optimal use of light.
Some species have irregular, or even branched palisade parenchyma cells (Y-shaped: arm palisade cells). Especially ferns, conifers and a few angiosperms (some Ranunculacea- and Caprifoliacea-species like elder) harbor such exceptions. No direct relations between these species exist. Does the branched palisade parenchyma have any selective advantage? It is suggested that the resulting enlargement of the surface is advantageous, but then again: why is this structure so rare?
The development and particularly the differentiation of the palisade parenchyma is influenced by extern factors like light and the CO2-content of the atmosphere. In many species, it is distinguished between sun- und shade leaves. Sun leaves have been exposed to large quantities of light during ontogenesis. This results in a multi-layered palisade parenchyma. Shade leaves in contrast perceive only little light, the palisade parenchyma stays single-layered. The enlargement of the palisade parenchyma causes usually a reduction of the spongy parenchyma, which is accordingly less well-developed in sun leaves. As important as the exposition to light is the leaf's position at the stem. Old leaves that live near the soil do often have a palisade parenchyma of just one layer's thickness, while younger ones at the top of the plant have normally multi-layered palisade parenchymas. This organization is most common, but the following variations can also be found:
|
|
palisade parenchyma at the undersurface of the leaf. It is especially striking in scales. Examples: arbor vitae (Thuja), leaves of ramsons (Allium ursinum). |
|
|
palisade parenchyma at both surfaces. This is typical for plants of dry habitats (xerophytes). Example: prickly lettuce (Lactuca serriola). |
|
|
ring-shaped, closed palisade parenchyma: occurs at cylindrically organized leaves and needles of conifers. |
The variability of the cells of the spongy parenchyma and that of its organization is even greater than that of the palisade parenchyma. It is often said to be an aerenchyma, since it is characterized by a large number of connected intercellular spaces. This does not mean that the contact between palisade parenchyma and intercellular spaces is less well developed. On the contrary: the proportion of palisade parenchyma that is in contact with the intercellular space is larger than that of the spongy parenchyma. The elongated and cylindrical shape of the cells enables only selective contact to neighboring cells even at close packing. The spongy parenchyma shows larger contact areas as well as cell-to-cell contacts between palisade parenchyma and spongy parenchyma and between spongy parenchyma and cells of the vascular bundles. This is also the reason for a water and assimilate transport that proceeds without losses.
The intercellular system causes the development of large inner surfaces within the leaf. The ratio of intercellular space to total cell volume is species- and habitat-specific. It ranges around 70-700:1000. F. M. TURREL gained the following results, when measuring and extrapolating the total foliage of a Catalpa in 1934:
![]() inner surface: 5100 m2
inner surface: 5100 m2
![]() outer surface: 390 m2
outer surface: 390 m2
The ratio of these two surfaces is again dependent on species and habitat. It is low in shade leaves and high in sun leaves.
Step by step, the two mesophyll types developed during evolution. Both spring from the same ground tissue and numerous transitions prove that the typical palisade and spongy parenchyma cells are merely extremes of a completed differentiation. With few exceptions, all cells of algae are able to perform photosynthesis. The leaves of simple bryophytes are generally single- and only rarely double-layered. No distinction can be made as to epidermis and mesophyll.
Other bryophytes, like the Polytrichum -species (mosses) and Riccia-species (liverworts) have lamellar ribs that consist of several layers of tissue on top of each other and are specialized for assimilation. Some liverworts (like Marchantia) have a multi-layered thallus that lives directly at the soil surface. Upon a layer of voluminous parenchyma cells sits an assimilation tissue of numerous small, plump cells gathered together in groups and interspersed with large, air-filled cavities. Liverworts have neither stomata nor guard mechanisms. Instead, their upper surface is mingled with characteristic pores that cannot be closed.
Many aquatic and marsh plants have a homogeneous aerenchym with very large intercellular cavities. They are not structured into palisade parenchyma and spongy parenchyma. Neither can it be distinguished between mesophyll and epidermis. The leaf tissue of the canadian waterweed (Elodea canadensis) is reduced to only two cell layers.
The vascular bundles of nearly all vascular plants are surrounded by a bundle sheath, a more or less extensive layer of parenchyma cells. Its cells contain usually less chloroplasts than those of other mesophyll cells. They are frequently also starch-deposits.
The cells of the bundle sheath of many -especially tropical- graminaceous plants are enclosed by a wreath of chloroplast-containing mesophyll cells (called Kranz anatomy; Kranz (german) = wreath). Both cell layers surround the vascular bundles -if seen in cross-section- like two concentric rings. The last years showed that this construction is typical for C4-plants. C4-plants are a group of species belonging to different mono- and dicot-families. They integrate carbon dioxide into different organic acids before advancing to photosynthesis. Plants lacking this metabolic pathway are termed C3-plants. Even within one and the same genus, one species may be characterized as a C4-plant while another is a C3-plant. The best known example are the two Atriplex-species A. patula (C3) and A. rosea (C4).
There are even some species where the lower leaves are constructed in one way and the upper ones in the other. Accordingly, they have different photosynthesis efficiencies. These species are usually succulent plants living in dry habitats. The stomata are closed during the day to avoid water loss. Hence, CO2 can only be taken up and accumulated at night (CAM-plants). Fixation is as in C4-plants; the respective organic acid (normally malate) is stored in the vacuoles of voluminous parenchyma cells lacking chloroplasts. During the course of the day, the carbon dioxide is split off again and, after diffusion into mesophyll cells, fed into photosynthetic pathways.
Leaves develop from leaf buttresses, that stem from small lateral bulges of the shoot. The flat structure of the leaf is caused by a preferred plane of division. This type of meristematic activity is attributed to the lamellar meristem, the predecessor of the mesophyll. Differentiation into the typical palisade parenchyma and the spongy parenchyma takes place due to the uneven growth rates of the different cell layers in the developing leaf. The activities of cell division and cell elongation occur in the different layers at different times. Additionally, the cells expand into different directions. An example: the cells of the palisade parenchyma develop by anticlinal division (at right angles to the leaf surface), those of the vascular tissue in parallel. As long as division and elongation of two tissues are concerted, the relation of the cells remains the same. But the cells of the epidermis start rather early during ontogenesis to divide more rapidly than those of the not fully differentiated parenchyma. This causes splaying of the parenchyma cells, whereby the intercellular spaces are generated. They are partially filled again upon further anticlinal division of the palisade parenchyma.
The differentiation of the lower epidermis and the spongy parenchyma is coordinated somewhat more variable. Depending on the species is either the cell division of the epidermis or that of the spongy parenchyma stopped first. Cell division and cell elongation are temporally separated from each other. The different growth rates of the different parts of the developing leaf succeed in a wave-like growth movement. These growth movements were documented in a time-lapse film. It shows impressively how the growth and expansion of a beech leaf spread in intensive wavy movements (W. KAUSCH, Universität Bonn, 1986; production and distributor: Institut für Wissenschaftlichen Film, Göttingen).