Before we deal with the architectural design of flowers in more
detail, it should be said that they are structures serving the sexual
reproduction: their final product is seed. Flowering plants are thus
also called spermatophytes (seed
plants). They include two large groups, the
gymnosperms and the
angiosperms. The gymnosperms are
the evolutionary older and more primitive group. They are, in a
strictly systematical sense, no class on their own but enclose two
real classes, the conifers and the
cycadeans that do not have the same
ancestors. The angiosperms that are
usually seen as the original flowering plants do all belong to just
one class. They represent the overwhelming majority of species and
what you will learn in the following is principally true for their
flowers.
It is taken for granted that everybody has through his own
experience gained an impression of the diversity of flower shapes,
colors and smells. It will be equally well known that these features
developed in an evolutionary process of mutual adjustments of flowers
and their individual pollinators (insects: bees, bumble-bees,
butterflies, flies, etc.; birds: hummingbirds, etc.). Plants that are
pollinated via wind have very unobtrusive flowers.
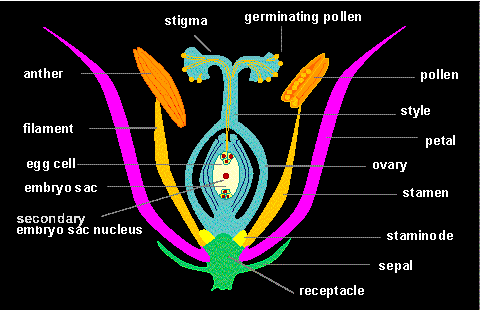
Diagram (cross section) of a
flower green: receptacle and sepals,
lilac: petals, orange: stamina, yellow:
staminods, blue: carpels
Flowers are short shoots with limited growth. The leaf organs are
arranged in several circles (whorls), one above the other at an often
strongly compressed flower axis. These structures differ strongly
both in their appearance and in their function from normal
leaves.

Progressing from bottom to top and from the out- to the inside can
it be distinguished between the usually unobtrusively colored
calyx that consists of sepals,
the conspicuously colored corolla
that is built of petals, the stamens and finally the carpels. Calyx
and corolla together are the
perianth. But if calyx and
corolla cannot be distinguished or if the calyx is missing it is
spoken of an incomplete flower or
perigon. Flowers with a missing
perianth are typical for species with wind-mediated pollination. The
totality of the stamens, the male reproductive organs, is called
androecium that of the female
reproductive organs is named
gynoecium.
The flower axis has hardly any appearance in most angiosperms. It
is usually shortened and broadened to the so-called
receptacle that is sometimes
slightly vaulted or disc-shaped. In gymnosperms, however, as well as
in some primitive angiosperms (Magnolia,
for example), it has the shape of a cone.
The flowers of angiosperms have originally only had an incomplete
perianth that can very likely be traced back to leaflike bracts and
occurs in an even more simplified version already in gymnosperms. The
majority of angiosperm flowers has a double perianth with a clear
separation of function between calyx and corolla. While the corolla
has as a rule the function of attracting pollinators (it is, after
all, designed mainly on insect aesthetics), the calyx is made to
protect the growing bud. It loses its function as soon as the bud
flowers; with some species it is even dropped off during flowering,
though with most species it just stops growing or shrivels. Only
rarely are the sepals reactivated after pollination. If they are, a
new growth starts whereby an involucre of the fruit is formed.
 All
flower structures have originally been organized in a tight screw
(Anemone). These flowers are strictly spoken asymmetric but
are nevertheless grouped together in books on classification with
those of a radial symmetry. They are
characterized by the arrangement of their flowers in whorls
(flowering circles). All structures of one whorl are of the same
appearance and every whorl has the same number of structures (three,
four and five are very common).
All
flower structures have originally been organized in a tight screw
(Anemone). These flowers are strictly spoken asymmetric but
are nevertheless grouped together in books on classification with
those of a radial symmetry. They are
characterized by the arrangement of their flowers in whorls
(flowering circles). All structures of one whorl are of the same
appearance and every whorl has the same number of structures (three,
four and five are very common).
Subsequent whorls are usually organized in a way that brings the
structures of one whorl in the gap positions of the preceding (rule
of alternation). Such flowers have two or more planes of symmetry;
sometimes those with only two planes are called bilateral. Even if
the gynoecium shows another number of organs, the flowers are grouped
together with those with radial symmetry.

Flowers that have only one plane of symmetry due to either
differing shapes of the structures of one whorl or due to incomplete
whorls are widespread. They are thought to be of a very high
developmental state and are called zygomorph
(monosymmetric or dorsiventral). Examples are the pansy (Viola)
or the labiates (Lamiaceae).

With most flowering plants, the petals are separate from each
other (dialypetalae) while they are fused to a tubular corolla with
sympetalae. The number of free points shows how many petals are
involved.
Both symmetry and architecture of the flower are clearest
represented via diagrams of the flowers. They show schematized
outlines where the single structures, their numbers and positions are
given. If such diagrams are compared to each other, it shows that
five is a typical number for the whorls of dicots while monocots have
mostly three organs per whorl.
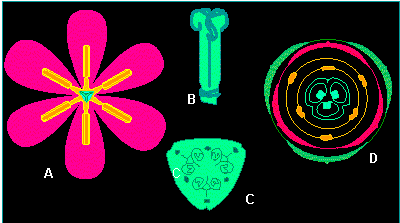
Tulipa gesneriana (tulip).
A. top view of flower,
B. and
C. pistil overall view,
B. cross section through the
ovary; D. generalized flower
diagram. Flower diagrams were drawn in the last century by the
German botanist A. EICHLER (1875, 1878) for most of the indigenous
and foreign plant groups. They symbolize the position, number and
the symmetry ratios of the flower's single components in every
circle of petals (according to W. TROLL, 1975).
The petals can often be traced back to reorganized stamens. This
interpretation is proven by the fact that links between both
categories of structures exist. A classic example is the white
waterlily (Nymphaea alba, a rather primitive angiosperm) but
such metamorphosis can, too, be found in many other species.
Cultivators use this feature to select filled flowers. The wild rose
(Rosa canina), for example, has only five petals and many
stamens while most of the cultured varieties are characterized by
filled flowers with many petals and a reduced number of fertile
stamens.
A normal stamen consists of a pollen-containing, fertile anther (a
group of two to four microsporangia) borne at the tip of a blade
stalk or filament. The anther has (usually) two theces that are
combined via the connective. Each theca includes two pollen
sacs where the production of the
pollen takes place.
Sterile stamens are called
staminodes. One variation are the
nectar-secreting honey leaves that live mostly between perianth and
stamens. They can adopt the size and color of petals and belong to
the pollinator-attracting apparatus of the plant, like in buttercup
(Ranunculus, Ranunculaceae).
The central part of a typical angiosperm flower is the
gynoecium that is composed of one
(simple pistil) or more (compound pistil)
carpels. The carpels are,
contrary to those of the gymnosperms, fused (see beneath). Their
original structure that of a leaf, is hardly recognizable. The
gynoecium contains one or more pistils.
A characteristic pistil has,
from bottom to top, an enlarged basal
ovary that includes the
embryo sac, a columnar
style and distal
stigma, the organ that receives
the pollen. Stigmata may be very varied in shape and structure. They
are often button-shaped and equipped with papills. It is not rare
that they are branched. It is distinguished between dry and moist
stigmata. The surface of the dry ones is often studded with
hair-shaped papills while moist ones are coated with a sticky film.
The style serves to bring the stigma into a favorable position for
pollination. The leafy character of the carpels can easily be seen
with a cross-section through the ovary. Both rims of a carpel seem to
be rolled in and fused so that a tube-shaped structure is formed. On
the basis of the thus built chamber (locule) are the seeds. This
protection is missing in gymnosperms. Here the seeds lie openly on
the carpels.
The suture that is formed by the fusion of the rims is called the
ventral suture in angiosperm
carpels while the midrib of the carpel is somewhat misleadingly
called the dorsal suture.
Normally the gynoecium of angiosperms is composed of several
carpels. A remarkable exception is made by the leguminoses where the
flower contains just one carpel. Each carpel of a gynoecium may
either form a pistil of its own
(choricarpy or
apocarpy) or several carpels may
be fused together to one pistil
(coenocarpy). The advantage of
the latter solution is in the ability to distribute the pollen tubes
among all carpels after successful pollination.
The way in which carpels are fused together and the resulting
position of the ovules within the ovary, the so-called
placentation, has been recognized
to be an important taxonomical feature for the classification of
angiosperms by A. ENGLER and E. PRANTL at the end of the last
century. Some orders have been named after their moulding of ovary
architecture, like for example centrospermae
or parietales.
An ovary that is built from free carpels is called
apocarp and is supposed to be the
most original form of an angiosperm gynoecium. It is often found in
ranunculaceae. Ovaries with fused carpels are derived from it.
In a syncarp ovary, the carpels are fused laterally. Its number of
locules fits that of its carpels that are separated from each other
by a septum. This can be seen very clearly with the ovary of a tulip.
The ovules are inserted at the inner angles of the locules (axial
placentation). False septums may have been put in later on which
could lead to the misapprehension of a syncarpous ovary at
superficial examination. But the unchanged parietal placentation of
the ovules enables a definite classification.
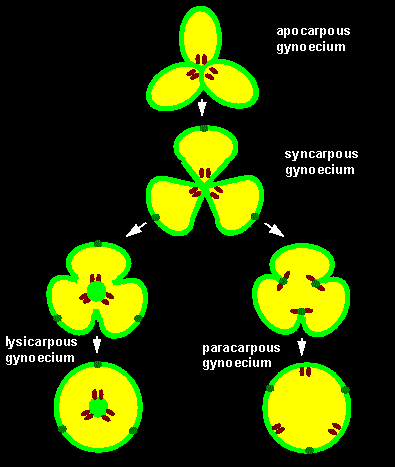
Scheme of evolution for the basic type of gynoecium. The
syncarpous gynoecium developed from the apocarpous one (on top)
which again differentiated into the paracarpous (right) and
lysicarpous (left) gynoecium. The paracarpous and the lysicarpous
types are depicted in two different stages of evolution to
elucidate the different ways of development from the basic
syncarpous type. The ovules are given in dark brown (according to
A. TAKHTAJAN, 1942).
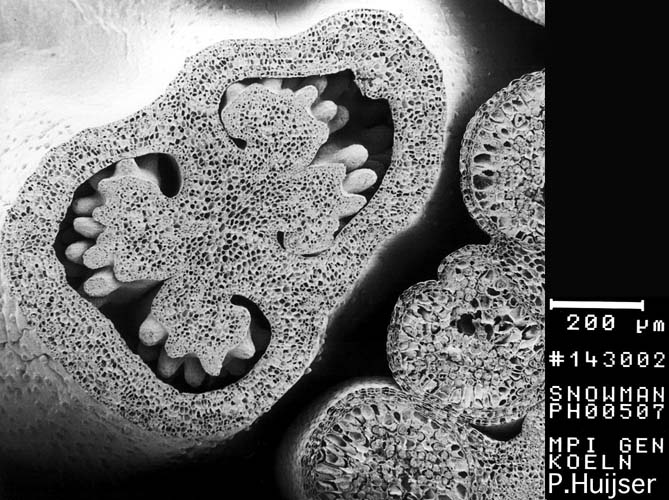
Scanning Electron Micropgraph: cross-section
through immature flower bud. Detail: Bilocular ovary with immature
ovules, anther with immature pollen (tetrad state) - Snapdragon
(Antirrhinum majus))
from
"Snapdragon" © K. STÜBER
Lysicarpous ovaries display a unified cavity formed through
disappearance of the lateral rims. Their ovules are inserted at an
axial cone in the centre of the ovary (free central placenta).
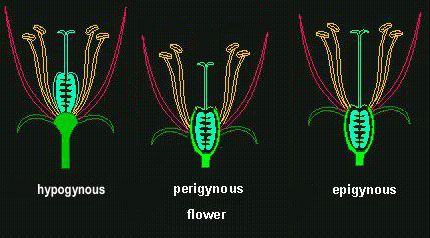 The position of the ovary in relation to the other structures of a
flower is an important taxonomical feature. It has to be
distinguished between a hypogynous (the perianth is attached to the
receptacle below the pistil), perigynous (perianth and stamens
are borne on the rim of a concave structure in the depression of
which the pistil is borne) or epigynous ovary (blossom seems
to arise upon or above the ovary).
The position of the ovary in relation to the other structures of a
flower is an important taxonomical feature. It has to be
distinguished between a hypogynous (the perianth is attached to the
receptacle below the pistil), perigynous (perianth and stamens
are borne on the rim of a concave structure in the depression of
which the pistil is borne) or epigynous ovary (blossom seems
to arise upon or above the ovary).
Flowers that contain both an androecium and a gynoecium are called
androgynous or
hermaphroditic. The flower is
male, if just one androecium is present; if only one gynoecium is
developed, then it is a female flower. A plant that has both male and
female flowers is called
monoecious while species where
male and female flowers live on different plants, are called
dioecious. In the extreme, the
flower is reduced to only one stamen or carpel. An especially
interesting case is the spurge (Euphorbia)
and some of its closer relatives. With them several extremely reduced
flowers are grouped together to a functionally hermaphroditic unity,
a cyathium. By integration of a
set of bracts that bear great, often yellowish nectaries and some
additional colored bracts, the impression of a normal hermaphroditic
flower is perfected (example: poinsettia;
Poinsettia pulcherrima).
© Peter v. Sengbusch - Impressum


