Botanik online 1996-2004. Die Seiten werden nicht mehr bearbeitet, sie bleiben als historisches Dokument der botanischen Wissenschaft online erhalten!
"Der Substanzverlust, der durch die Atmung nebenbei erzeugt wird, hat den Zweck, mechanische Kräfte zu entwickeln, durch welche die Atome und Moleküle der übrigen Substanzen in diejenigen Bewegungen versetzt werden, aus denen Wachstum und die sonstigen Funktionen der lebenden Pflanze resultieren. Mit einem Wort: Die Atmung ist die Kraftquelle, aus der alle Lebenserscheinungen ihre Kräfte schöpfen, während die Assimilation in den chlorophyllhaltigen Organen die Stoffe liefert, die später zum Zweck des Lebens in Bewegung gesetzt werden" (J. v. SACHS. 1882)
Alle Reaktionsschritte der Glykolyse laufen im Cytosol der Zellen ab. Der Citratzyklus und die Atmungskette sind räumlich von ihnen getrennt. Die Atmungskette stellt einen membrangebundenen Vorgang dar, der an die innere mitochondriale Membran (Sacculusmembran) gebunden ist. Mitochondrien werden üblicherweise als die Kraftwerke der Zelle bezeichnet, weil in ihnen unter Sauerstoffverbrauch ATP gebildet wird. Der Vorgang wird auch oxydative Phosphorylierung genannt. Er ähnelt in manchem der Photophosphorylierung während der Photosynthese: Beide Prozesse funktionieren nur dann, wenn intakte Membranen vorhanden sind, die sich ihrerseits zu Vesikeln schließen und so ein Kompartiment der Zelle von einem anderen trennen.
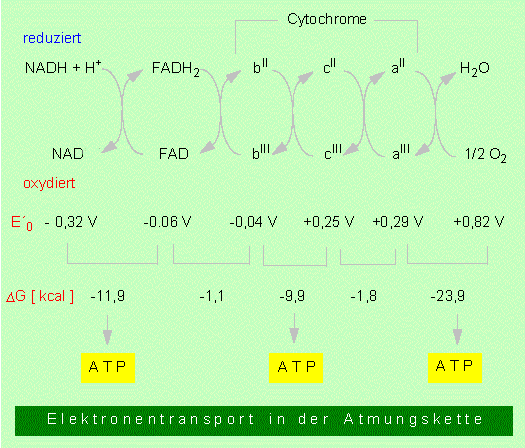
Zusammenfassend kann die Atmungskette als der Prozeß charakterisiert werden, bei dem NADH + H+ und FADH2 unter Sauerstoffverbrauch oxydiert werden, Wasser entsteht und gleichzeitig ATP gebildet wird. Bekanntlich ist eine Wasserbildung (Knallgasreaktion)
2 H2 + O2 > 2 H2O
stark exergonisch. Die Atmungskette führt zwar im Endeffekt zum gleichen Ergebnis, nur läuft die Reaktion über eine Anzahl von Zwischenstufen (eine Elektronentransportkette), bei der die Energie sukzessive freigesetzt und in energiereiche Bindungen investiert wird.
1. NADH + H+ wird oxydiert, es entsteht NAD+. Der Wasserstoff wird von einem weiteren Wasserstoffakzeptor, dem FAD übernommen.
2. FADH2 wird durch Cytochrome oxydiert. Wie schon früher ausgeführt, handelt es sich hierbei um Proteine (Cytochrom b, Cytochrom c, Cytochrom a u.a.), die als Kofaktoren Hämgruppen (Porphyrinringe) tragen. In ihrem Zentrum befindet sich ein Eisenion mit wechselnder Valenz:
FeIII < > FeII
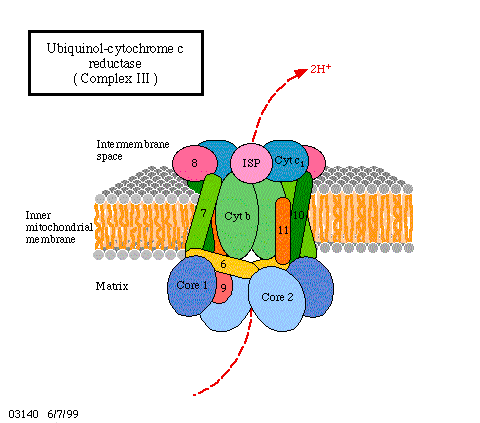
Während das FAD außer den Elektronen auch noch die Protonen trägt, trifft das auf die Cytochrome nicht mehr zu, denn sie übertragen nur Elektronen. Die Protonen werden in Lösung entlassen. In der Atmungskette wird das FeIII des Cytochroms b zunächst in FeII überführt. In pflanzlichen Zellen sind mindestens drei verschiedene Cytochrom-b-Typen in Serie hintereinandergeschaltet. Die Rückführung in FeIII erfolgt durch Elektronenabgabe an das Cytochrom c, das die Elektronen in einem weiteren Schritt an Cytochrom a weiterreicht. Zwischen dem Flavoprotein und den Cytochromen kann ein Chinon/Hydrochinonsystem zwischengeschaltet sein.
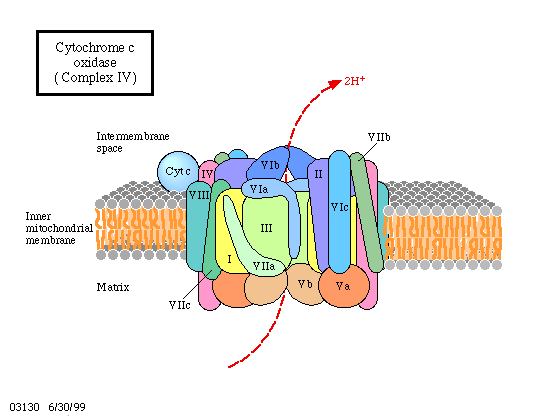
3. Das reduzierte Cytochrom a gibt die Elektronen an Sauerstoff ab, der sich dann umgehend mit freien Protonen zu Wasser verbindet.
Bei der Besprechung der Redox-Reaktionen wird gezeigt, daß jedes Redoxpaar durch ein Redoxpotential (E'°) charakterisiert und daß dieses wiederum der freien Energie der Reaktion (delta G°) proportional ist. Die für die Atmungskette ermittelten Werte sind der Abbildung zu entnehmen. Man erkennt hier, daß bei drei Redoxpaaren die Potentialdifferenz ausreicht, um je ein ATP-Molekül zu formen. Folglich entspricht ein NADH + H+ -Äquivalent drei ATP-Molekülen, während ein FADH2 lediglich zur Bildung von zwei ATPs führt.
Wie kann man sich das alles mechanistisch, molekularbiologisch vorstellen? Dazu muß einmal erinnert werden, daß alle genannten Kofaktoren proteingebunden sind, daß diese Proteine entweder integrale Bestandteile der Membran oder eng mit ihr assoziiert sind. Es ist darüber hinaus wichtig, zu wissen, daß die Orientierung der Proteinmoleküle in der Membran stets gleich (vektoriell) ist und daß diese Membran protonenundurchlässig ist. Außer den genannten Proteinen wird noch ein weiteres Enzym, eine ATP-Synthetase, benötigt. Wie sind nun ATP-Synthetaseaktivität und Elektronentransportkette miteinander gekoppelt? Es gab schon seit langem gute Hinweise darauf, daß dieses Enzym selbst die Funktion eines Kopplungsfaktors wahrnimmt.
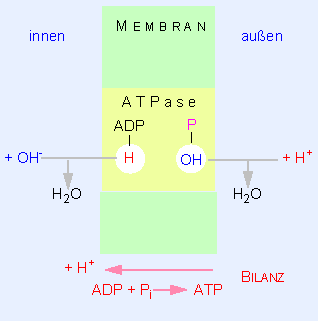
Ein Modell des Mechanismus der Protonentranslokation durch die innere mitochondriale Membran. Man beachte, daß nicht Protonen wandern, sondern daß bei der ATP-Synthese auf der einen Membranseite (außen) OH-Ionen, auf der entgegengesetzten (innen, Matrixseite) Protonen freigesetzt werden. Die Bilanz der Reaktion führt zu einer scheinbar gerichteten Protonenwanderung. Die ATPase verfügt über getrennte, aber benachbarte Bindungsorte für ADP und Phosphat.
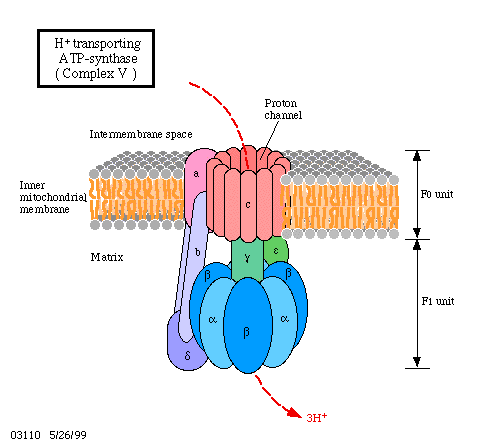
1961 postulierte P. MITCHELL, daß Protonen während des Ablaufs der Atmungskettenreaktionen gerichtet durch die Membran befördert werden. Damit baut sich ein pH-Gradient auf. Der "Rücktransport" erfolgt über einen spezifischen Protonenkanal. Mit diesem Rücktransport ist eine ATP-Synthese gekoppelt, bei der die potentielle Energie des Protonengradienten in die dritte Phosphatbindung des ATP einfließt.
Die Beschreibung der Atmungskette sollte nicht zu dem Schluß verleiten, daß jetzt tatsächlich ganz bestimmte Protonen und Elektronen von einem Träger zum nächsten weitergereicht werden, vielmehr handelt es sich um Nettobilanzen, die den Zustand und das Gleichgewicht der jeweiligen Akzeptoren beschreiben.
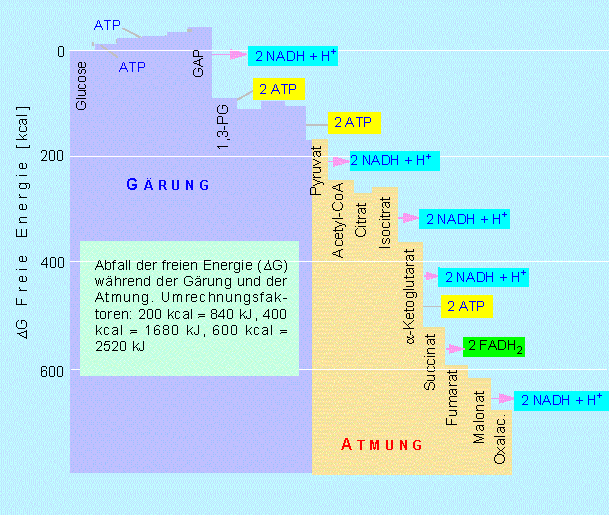
Abschließend können wir eine Gesamtenergiebilanz des Glucoseabbaus aufstellen: Die Oxydation ist mit einem Abfall der freien Energie verbunden; bei der vollständigen Verbrennung von Glucose fallen 686 kcal/mol (ca. = 2881 kJ/mol) an. Wieviel von diesem Energiebetrag kann die Zelle für ihren eigenen Bedarf nutzen?
Es werden pro Mol Glucose sechs Mol ATP gebildet (Substratkettenphosphorylierung). Die Zahl ergibt sich aus 3 x 2, da ja alle Schritte nach der Fructose-1,6-bisphosphatspaltung doppelt zu zählen sind. Von diesen sechs ATPs werden für den Start der Glykolyse zwei verbraucht, es verbleiben vier.
Während der Glykolyse bis zur Stufe des Acetyl-CoA werden 2 x 2 NADH + H+ gebildet. Im Citratzyklus entstehen 3 x 2 NADH + H+ und 1 x 2 FADH2. Über die Atmungskette verrechnet, bedeutet das einen Gewinn von 34 ATP-Äquivalenten. Das ergibt in der Summe 38 Mole ATP pro Mol Glucose. Da in jeder energiereichen Bindung des ATP -7,3 kcal/mol (= ca. -30,6 kJ/mol) gespeichert sind, erhalten wir insgesamt 277 kcal/mol (ca. 1163 kJ/mol). Das wiederum entspricht, bezogen auf 686 kcal/mol, 40,6 Prozent der theoretisch möglichen Ausbeute. Die restlichen 59,4 Prozent werden als Wärme freigesetzt.
Mit dem Ergebnis sollte man zufrieden sein, wenn man bedenkt, daß die Ausbeute technischer Maschinen (Dampfmaschine, Benzinmotor) bei oder unter 20 Prozent liegt.