Botanik online 1996-2004. Die Seiten werden nicht mehr bearbeitet, sie bleiben als historisches Dokument der botanischen Wissenschaft online erhalten!
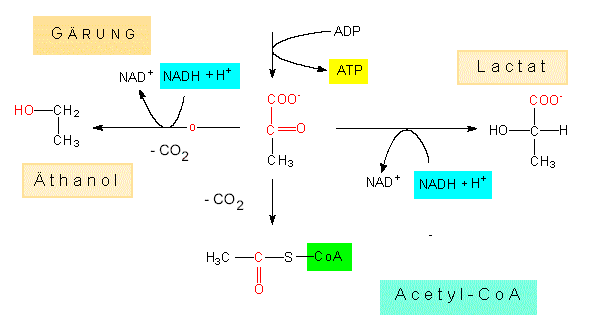
Die Glykolyse repräsentiert einen bei aerob und bei anaerob lebenden Organismen weit verbreiteten Abbauweg der Glucose. Andere Zucker und Polysaccharide müssen zunächst in sie oder eines ihrer phosphorylierten Derivate überführt werden. Im Verlauf der Degradation wird ATP gebildet (Substratkettenphosphorylierung). Rein formal kann Pyruvat (das Salz der Brenztraubensäure) als vorläufiges Endprodukt des Abbaus angesehen werden, denn hier trennen sich die Wege: Unter anaeroben Bedingungen wird es hydriert, wobei je nach Organismenart Lactat (z.B. in Milchsäurebakterien) oder Äthylalkohol (z.B. bei Hefe) entstehen. Führt die Glykolyse zu Endprodukten dieser Art, spricht man von Gärung. Unter aeroben Bedingungen wird es über ein Zwischenprodukt in den Citratzyklus eingeschleust, und über eine Elektronentransportkette (Atmungskette) wird ein ansehnlicher Energiebetrag in Form von ATP gewonnen. Die Ausgangssubstanz Glucose wird dabei vollständig zu Wasser und Kohlendioxyd oxydiert.
Im Jahre 1905 beobachteten A. HARDEN und W. YOUNG, daß der Glucoseabbau zum Erliegen kommt, sofern nicht ausreichende Mengen an anorganischem Phosphat angeboten werden. Es wird letztlich dazu benötigt, Zucker zu phosphorylieren. Sie isolierten ein Hexosediphosphat, das später als Fructose-1,6-diphosphat identifiziert wurde, und zeigten, daß es sich dabei um ein Zwischenprodukt des Glucoseabbaus handelt.
Der Abbauweg wurde in den Jahren vor 1940 restlos aufgeklärt. Die Biochemiker G. EMBDEN, O. MEYERHOFF, C. NEUBER, J. PARNASS, O. WARBURG, G. und C. CORI hatten den Hauptanteil am Erfolg.
Die Glykolyse ist auch unter der Bezeichnung
EMBDEN-MEYERHOFF-PARNASS-Schema in die Literatur eingegangen.
Die Reaktionsschritte sind wie folgt zusammenzufassen.
Die Reaktionskette beginnt mit einem irreversiblen Schritt, nämlich der Phosphorylierung der Glucose unter ATP-Verbrauch. Im zweiten - reversiblen - Schritt, wird das gebildete Glucose-6-Phosphat zu Fructose-6-Phosphat isomerisiert.
|
Unter Verbrauch eines zweiten ATP-Moleküls entsteht Fructose-1,6-diphosphat. Auch dieser Schritt ist nicht umkehrbar, denn das delta G° des phosphorylierten Zuckers reicht auch hier nicht aus, um die Phosphatgruppe auf ADP zurück zu übertragen. Es sind im Verlauf der gesamten Glykolyse die einzigen Reaktionen, die vektoriell (gerichtet) ablaufen. Schreibt man das Fructose-1,6-diphosphat (= Fructose-1,6-bisphosphat - FDP) in linearer Form, erkennt man, daß die beiden Phosphatgruppen terminal sitzen und dadurch die mittlere -C-C-Bindung so schwächen (Dipole in beide Richtungen), so daß das Molekül Zerfallstendenzen zeigt. Unter Mitwirkung des Enzyms Aldolase entstehen als Spaltproduke D- Glycerinaldehyd-3-Phosphat (GAP) und Dihydroxyacetonphosphat (DAP), die reversibel ineinander überführt werden können. Unter physiologischen Bedingungen liegt das Gleichgewicht weit auf der Seite des Dihydroxyacetonphosphats. Da Glycerinaldehyd-3-Phosphat durch nachfolgende Reaktionen kontinuierlich abgezogen wird, kann es aus dem Pool des Dihydroxyacetonphosphats ständig nachgeliefert werden.
Jetzt folgt der erste Oxydationsschritt, obwohl kein Sauerstoff im Spiel ist. Durch Dehydrierung freiwerdende Protonen und Elektronen werden von NAD+ übernommen. Die Aldehydgruppe des Glycerinaldehyd-3-Phosphats wird zu einer Karboxylsäureestergruppe des Phosphats, wozu freies anorganisches Phosphat benötigt wird.
Das entstehende 1,3- Diphosphat-D-Glycerat ist extrem labil und verliert leicht eine seiner Phosphatgruppen, die ihrerseits auf ADP übertragen wird. Damit ist unser Einsatz von vorhin wieder hereingeholt. Zwei Moleküle ATP wurden investiert, und zwei kommen wieder heraus.
Warum zwei? Aus dem ursprünglichen C6-Körper (einem Molekül, das sechs C-Atome enthält) sind nach Spaltung des Fructose-1,6-diphosphats zwei C3-Körper entstanden, und das wiederum heißt, daß wir die Gewinn- und Verlustbetrachtungen von da ab verdoppeln müssen.
Die beiden folgenden Schritte sind vielleicht weniger interessant, wichtig hingegen ist wieder die Dephosphorylierung des Phosphoenolpyruvats unter ATP-Bildung zu Pyruvat. An dieser Stelle kann die Besprechung der Glykolyse unterbrochen werden. Wie schon eingangs erwähnt, können von hier ab unterschiedliche Abbauwege eingeschlagen werden; z.B. entsteht, unter Mitwirkung des Enzyms Lactatdehydrogenase und unter NADH + H+ -Verbrauch Lactat, in anaerob wachsenden Hefezellen unter Mitwirkung von Pyruvatdecarboxylase, von Alkoholdehydrogenase und wiederum von NADP + H+, Äthanol. Der Nettogewinn bei diesen Prozessen (Gärungen) ist recht bescheiden, denn er beruht nur auf zwei bereits genannten ATP-Molekülen. Unter aeroben Bedingungen läuft die Reaktionskette vom Pyruvat zum Acetyl-CoA:
Pyruvat + NAD+ + CoA > Acetyl-CoA + CO2 + NADH + H+ .
Hierbei handelt es sich um eine oxydative Decarboxylierung, weil
Kohlendioxyd freigesetzt und das Pyruvat gleichzeitig dehydriert
(oxydiert) wird. Hierbei spielt das Thiaminpyrophosphat, das Koenzym
der Pyruvatdehydrogenase eine entscheidende Rolle.
| Acetyl-CoA C N O P S |
Die Schlüsselrolle zum Einstieg in den Citratzyklus spielt das Acetyl-CoA.
Das Koenzym A (CoA) ist ebenfalls ein Kofaktor, also eine prosthetische Gruppe wie das NAD+ und die anderen bisher besprochenen. Die terminale SH-Gruppe bildet das reaktive Zentrum, das eine C2-Einheit (einen Acetylrest) binden und damit auf andere Molekülgruppen .übertragen kann. Vom Acetyl-CoA (aktivierte Essigsäure) zweigt einmal die Fettsäuresynthese ab, zum anderen kann die aktivierte Essigsäure unter Abspaltung des Koenzyms an Oxalacetat gekoppelt werden und damit sind wir bereits im Citratzyklus.
Über eine Reihe von Zwischenprodukten führt er zur Regeneration des Oxalacetats, womit die Benennung Zyklus gerechtfertigt ist.
Doch passiert etliches mehr:
An zwei Stellen der Reaktionskette wird Kohlendioxyd, eines der beiden Oxydationsprodukte der Glucose, abgespalten. Der in den Zyklus eingeschleuste C2-Körper ist damit vollständig oxydiert.
An vier Stellen werden je 2 H+ + 2 e- freigesetzt, an dreien werden sie auf NAD+ übertragen, an einer (bei der Dehydrierung von Succinat zu Fumarat) auf FAD.
- Einige der Zwischenprodukte sind Ausgangsstoffe von Biosynthesewegen.
Es wird ein Molekül ATP (bei Tieren: GTP) gebildet. Seiner Entstehung geht die Bildung von Succinyl-CoA voraus. Diese Verbindung enthält eine energiereiche C~S-CoA-Bindung (Thioesterbindung, delta Go: -8 kcal/mol = ca. -33,6 kJ/mol), deren Spaltung Energie für eine ATP (bzw. GTP)-Bildung bereit hält.
| > |