Botanik online 1996-2004. Die Seiten werden nicht mehr bearbeitet, sie bleiben als historisches Dokument der botanischen Wissenschaft online erhalten!
Der Wissenschaftszweig, der sich mit Geweben befaßt, ist die Histologie. Für die Beschreibung pflanzlicher Gewebe wird jedoch genauso oft der Ausdruck Pflanzenanatomie verwendet. Der Begriff Gewebe beruht eigentlich auf einer Fehlinterpretation, denn einer der ersten Pflanzenanatomen, der Engländer N. GREW, meinte (1682), daß das Zellwandgerüst der Pflanzen aus äußerst feinen Fäden bestehen würde und daß der Aufbau des Pflanzenkörpers mit einer Übereinanderschichtung einer größeren Anzahl Brüsseler Spitzen vergleichbar sei. In diesem Sinne sprach er von einem Zellgewebe (Contexus cellulosus). Trotz dises Mißverständnisses blieb der Name erhalten und wurde sogar von der Pflanzenanatomie auf die tierische Histologie übertragen.
So wie wir es heute sehen, sind Gewebe Zusammenschlüsse von Zellen; Organe sind funktionelle Einheiten eines Organismus. Die Grundorgane einer phanerogamen Pflanze sind Sproß, Blätter und Wurzel.
Organe bestehen aus unterschiedlichen Geweben, die Blätter z.B. aus Abschlußgewebe, Assimilationsgewebe, Leitgewebe u.a. Ein Gewebe wiederum kann unterschiedlich strukturierte Zelltypen enthalten. So enthält das Leitgewebe die Zellen des Xylems und die des Phloems.
Auf Vorschlag von C. W. v. NÄGELI unterscheidet man zwischen Teilungs- und Dauergeweben. Teilungsgewebe (Meristeme) verfügen über ein hohes Zellteilungsvermögen. In Dauergeweben kommt es nur ausnahmsweise zu Zellteilungen. Diese Gewebe bestehen in der Regel aus weitgehend ausdifferenzierten, oft spezialisierten Zellen. Aufgrund ihrer funktionellen Eigenschaften können sie folgenden Kategorien zugeordnet werden:
![]() Abschlußgewebe
(Epidermis, Kork, Borke)
Abschlußgewebe
(Epidermis, Kork, Borke)
![]() Grundgewebe (Parenchym)
Grundgewebe (Parenchym)
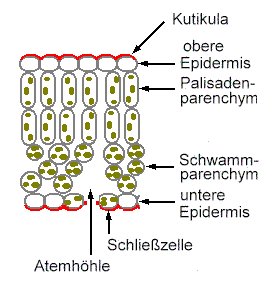
![]() Assimilationsgewebe
(Palisadenparenchym, Schwammparenchym)
Assimilationsgewebe
(Palisadenparenchym, Schwammparenchym)
![]() Festigungs- oder Stützgewebe (Kollenchym, Sklerenchym)
Festigungs- oder Stützgewebe (Kollenchym, Sklerenchym)
![]() Leitgewebe (Gefäße, Xylem, Phloem).
Leitgewebe (Gefäße, Xylem, Phloem).
1877 erschien A. de BARYs grundlegendes Werk über die Gewebelehre: "Vergleichende Anatomie der Vegetationsorgane der Phanerogamen und Farne", aus dem das folgende Zitat stammt:
"Die Gewebeelemente jeder Art gehen aus den Zellen des Meristems hervor, jedes hat ursprünglich die Eigenschaften einer Zelle. Mit der definitiven Ausbildung tritt nun zunächst der Hauptunterschied ein, daß die einen zeitlebens den Bau und alle charakteristischen Eigenschaften typischer Zellen behalten, die anderen die Zellqualitaten verlieren. Erstere behalten die Fähigkeit selbständigen Wachstums und bleiben teilungsfähig, als Folge davon kann aus ihnen selbst Meristem werden, die anderen verlieren mit ihrer Ausbildung die Fähigkeit der Zellteilung und des selbständigen Wachstums. In der Regel hören sie überhaupt auf, zu wachsen. In manchen Fällen findet ein andauerndes wirkliches Wachstum solcher Elemente in Folge ihrer Ernährung durch benachbarte Zellen statt"
 |
Mit den Ursachen des Differenzierungsprozesses werden wir uns an anderer Stelle befassen. Hier nur einige immer wiederkehrende Teilungsmuster, aus denen sich bereits eine ganze Menge über die Gestalt und die Größe der Zellen herleiten läßt:
Das wohl wichtigste Prinzip einer pflanzlichen Differenzierung ist die Polarität, die während der Embryonalentwicklung (Ontogenese) bereits nach der ersten Teilung der befruchteten Eizelle (der Zygote) zum Ausdruck kommt und die Hauptachse des Vegetationskörpers damit irreversibel festlegt. Wurzeln und Sproß entwickeln sich nachfolgend unabhängig voneinander. Man spricht daher von einer Wurzel-Sproß-Polarität.
Zellteilungen, die senkrecht zur nächsten Oberfläche des betreffenden Organs erfolgen, nennt man antiklin, solche, die parallel zu ihr angelegt sind, periklin. Oft teilen sich die Zellen inäqual und als Ergebnis erhält man zwei ungleich große Tochterzellen. Als Faustregel gilt, daß die kleinere den vorhandenen physiologischen Zustand beibehält, während sich die größere in eine bestimmte Richtung differenziert bzw. spezialisiert. Ausnahmen von dieser Regel kommen vor, so spezialisiert sich bei der Spaltöffnungsbildung die kleinere Tochterzelle stärker als die größere.
Zu den auffälligsten Merkmalen unterschiedlich differenzierter Zellen gehört die ungleiche Größenzunahme. Manche der Zellen teilen sich ohne eine merkliche Volumenzunahme, andere wiederum stellen die Teilungen ein und vergrößern sich beträchtlich. Pflanzliches Wachstum kann daher weitgehend auf eine Volumenzunahme einzelner Zellen zurückgeführt werden. Da nun aber der Vergrößerung einer jeden Zelle der Widerstand bereits vorhandener, benachbarter Zellen entgegenwirkt, die Zellen zudem auch noch über die Mittellamelle relativ fest untereinander verkittet sind, kommt es zwangsläufig zu beträchtlichen Gewebespannungen. Breitet sich eine Zelle in alle Richtungen gleichmäßig aus, würde man rein theoretisch eine Kugelgestalt erwarten. Da die Zellen meist aber von einer Vielzahl von Nachbarzellen umgeben sind, die wegen der Asynchronie der Zellteilungen und unterschiedlicher eigener Entwicklungsgeschichte verschieden groß sind, sind auch die zwischen den Zellen ausgebildeten gemeinsamen Wandflächen ungleich groß. Folglich ist die Form einer sich nach allen Seiten gleichmäßig vergrößernden Zelle als ein vielseitiger Körper (= isodiametrische Zelle, ein Polyeder) zu beschreiben. Oft erfolgt die Streckung der Zellen gerichtet. Meist geschieht das in Richtung der Achse des betreffenden Organs (Streckungswachstum). Die entstehenden Zellen sind langgestreckt und laufen an den Enden spitz aus. Für diesen Zelltyp wurde der Begriff prosenchymatisch geprägt.
Werden durch ein Streckungswachstum alle in einem bestimmten Gewebeabschnitt liegenden Zellen erfaßt (z.B. in den Wachstumszonen des Sprosses und der Wurzel), verlängert sich das Organ auf allen Seiten gleichmäßig, während das Wachstum nur einer Seite (Flanke) des Organs zu einer Krümmung führt. Im Zusammenhang mit den pflanzlichen Wachstumsbewegungen werden wir uns mit dieser Erscheinung noch ausgiebig auseinandersetzen.
Allein diese wenigen, durch ungleiche Teilungsraten und unterschiedliche Volumenzunahme einzelner Zellen verursachten Spannungen und Formveränderungen machen deutlich, daß der Kormus der Pflanze kein einheitlicher, durch einfache geometrische Beziehungen zu beschreibender Körper ist.
Die scheinbaren Unregelmäßigkeiten der Teilungen (unterschiedliche Teilungsaktivitäten einzelner Zellen oder Zellgruppen, inäquale Teilungen usw.), das unterschiedliche Streckungsvermögen einzelner Zellen und das Ausmaß sowie die Art ihrer Spezialisierung sind demnach die Ursache für die histologische Variabilität und die Entstehung einer spezifischen Gestalt der Pflanze (Morphogenese).
Obwohl jeder der Schritte, getrennt betrachtet, als eine Abweichung von der Norm erscheint, stehen sie zusammengenommen unter der Kontrolle des Genoms der Pflanze. Mit anderen Worten: Wachstum und Differenzierung sind exakt aufeinander abgestimmt und so koordiniert, daß die entstehende Pflanze eine artspezifische Gestalt annimmt. Es gibt zwischen den Zellen demnach einen Informationsaustausch, durch den deren Aktivitäten reguliert werden.
Phänologisch wird der Abgleich am eindrucksvollsten an der Ausbildung symmetrischer Formen auf zellulärer und auf Organebene sichtbar. Doch wie kann eine solche Kontrolle des Wachstums und die Koordination der Differenzierungsprozesse aussehen?
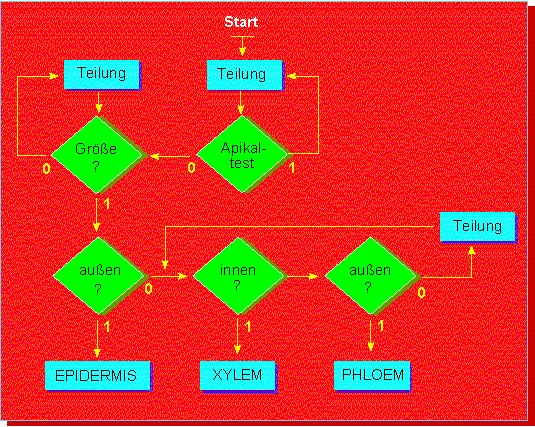
J. BONNER vom California Institute of Technology in Pasadena stellte
1965 ein Modell in Form eines Fließdiagramms vor, in dem das Zusammenspiel
genetisch fixierter Schaltstellen (Genen oder Gengruppen) für die
Entwicklung spezialisierter Zellen aus einer Einzelzelle (einer meristematischen
Zelle im Vegetationspunkt) dargelegt ist. Es mag uns immer noch zu hypothetisch
erscheinen, aber es stellt sicher eine sinnvolle Arbeitshypothese dar,
die dazu anregen soll, nach den geforderten Regulatoren zu suchen.