
Botanik online 1996-2004. Die Seiten werden nicht mehr bearbeitet, sie bleiben als historisches Dokument der botanischen Wissenschaft online erhalten!
http://www.mpiz-koeln.mpg.de/~saedler/Gentechnologie.html
Die im Neolithikum beginnende Domestizierung von Pflanzen war ein wichtiger Schritt in der Entwicklungsgeschichte der Menschheit. Hierdurch wurden die Grundlagen für die Entstehung humaner Hochkulturen gelegt. Die fortschreitende Anpassung der Pflanzen an die sich im Laufe der Zeiten ändernden Wünsche und Bedürfnisse der Menschen führte zu den heute bekannten Kulturpflanzen, deren überlebensfähigkeit in "freier" Natur - wenn überhaupt - nur sehr eingeschränkt möglich ist.
In fast allen Erdteilen wurden von den unterschiedlichsten Völkern Pflanzen domestiziert und, was wesentlich erscheint, zum Anbau in den folgenden Jahrhunderten über den ganzen Erdball verteilt. Diese so kultivierten Pflanzen wurden besonders auf hohe Erträge und gute Verdaulichkeit ausgewählt, was das Anwachsen der Erdbevölkerung auf gegenwärtig über 5 Milliarden Individuen ermöglichte.
Die Verteilung der Menschen (Abb.1) und ihre Wachstumsrate ist auf den einzelnen Kontinenten jedoch recht unterschiedlich.
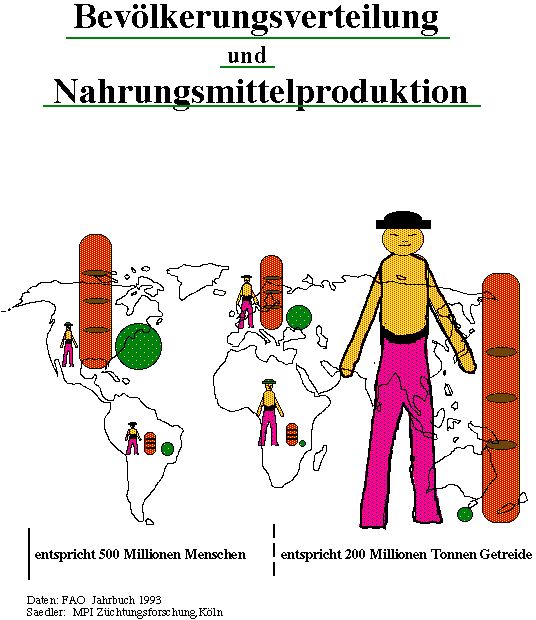
Bevölkerungsverteilung und Nahrungsmittelproduktion
Hieraus resultieren für jeden Kontinent spezifische Probleme, die hier nicht im einzelnen betrachtet werden können, vielmehr sollen nachfolgend drei Probleme angesprochen- werden, bei deren Lösung gentechnologisch veränderte Pflanzen einen Beitrag leisten könnten:
Wie die Abb.1 verdeutlicht, ist nicht nur die Bevölkerung auf die einzelnen Kontinente unterschiedlich verteilt, sondern auch die landwirtschaftliche Produktion in den Kontinenten recht verschieden. Lediglich in Nordamerika und Europa werden
Agrarüberschüsse erzielt, die u.a. zum Ausgleich von Defiziten in anderen Teilen der Welt eingesetzt werden.
Im asiatischen Raum leben mehr als 60% der Weltbevölkerung, deren vornehmlich auf Reis basierende Agrarproduktion für eine kalorische Versorgung nicht ausreichend ist. Ebenso defizitär ist die Versorgung mit Proteinen und Fetten in Afrika, Asien und Südamerika (Abb.2). Dies ist deswegen so besorgniserregend, weil dort die Bevölkerung immer noch exponentiell wächst, und so zukünftig ein immer größer werdender Nahrungsbedarf entsteht. Diese Spirale des Hungers kann nur bei einer sinnvollen Kontrolle des Bevölkerungswachstums durchbrochen werden .

Nahrung pro Kopf und Tag (1988-1990)
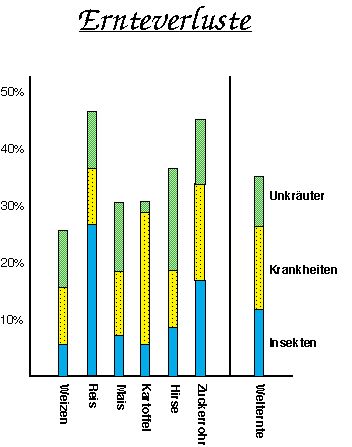
Ernteverluste
Andere große Verluste treten bei der Lagerung und dem Transport auf. Dies im einzelnen aufzuführen ist im Rahmen dieser Darstellung nicht möglich, bietet aber Ansatzpunkte zur Steigerung der Menge, der zur Verfügung stehenden Nahrungsmittel. Eine weitere Möglichkeit stellt die Herstellung von für den Anbau optimierten Nutzpflanzen dar, die nachfolgend erläutert werden soll.
Die Nutzung männlich steriler Linien in der Pflanzenzüchtung wurde durch den Erfolg der Hybridsaatzucht bei Mais seit Mitte der dreißiger Jahre offensichtlich. Mais hat gegenüber den meisten Gräsern den züchterischen Vorteil, daß die Geschlechter räumlich getrennt auf verschiedenen Blütenständen derselben Pflanze verteilt sind. Dies ermöglichte mit relativ bescheidenen Mitteln, reine Linien zu erstellen und anschließend durch Kreuzung zweier unterschiedlicher Inzuchtlinien Hybridlinien herzustellen, deren Erträge alle bisherigen Sorten weit übertrafen. Die Herstellung der Hybridlinien im großen Stil wird durch männliche Sterilität eines Partners erleichtert. Dies konnte bei Mais durch manuelle Entfernung der männlichen Blütenstände leicht erreicht werden. Das Verfahren wurde lange praktiziert bis infolge der Verteuerung menschlicher Arbeitskraft einerseits und der Entwicklung eines biologischen Sterilitätssystem andererseits Hybridsaatgut kostengünstiger hergestellt werden konnte. Mehrere biologische Systeme der cytoplasmatischen männlichen Sterilität waren beim Mais entdeckt worden. Allerdings nützen derartige sterile Systeme wenig, wenn sie nicht restauriert werden können. Neben der männlich sterilen Linie ist also auch eine "Restorer" Linie für die Entwicklung eines Hybridsaatzuchtprogrammes notwendig.
Die Erträge des Anbaus von Hybridsaatgut sind so hoch, daß bei optimalem Anbau und ausreichender Düngung in einem Zeitraum von etwa 50 Jahren eine mehr als 4-fache Ertragssteigerung (Abb.4) erzielt werden konnte.

Hybridsaatzucht
Die Tatsache, daß für die nächste Wachstumsperiode kein Saatgut zurückbehalten werden kann, beruht auf seinem hybriden Charakter, der keine Einheitlichkeit des Materials für einen weiteren Anbau gewährleistet. In anderen Worten: Hybridsaatgut muß ständig vom Züchter geliefert werden. Dennoch ist diese Strategie beim Mais so erfolgreich, daß eine Ausdehnung auf andere Kultursorten angestrebt wurde.
Obwohl in einigen Kulturpflanzen ebenfalls cytoplasmatisch männliche Linien bekannt waren, hatte das Desaster bei Mais im Jahre1970 in den USA den Optimismus zur Nutzung dieser Systeme zerstört. Wegen des einheitlichen Cytoplasmas des Hybridmaterials und aufgrund der Tatsache, daß ein Pilz sich in diesem Material besonders gut vermehren konnte, war durch eine explosionsartige Ausbreitung der "Northern Leaf Blight" ein Schaden von mehreren Milliarden Dollar entstanden. Cytoplasmatische männliche Sterilität wurde daher nicht weiter zur Herstellung von Hybridsaatgut beim Mais verwendet, vielmehr kehrte man zu alten Verfahren zurück, oder nutzt zunehmend moderne Wege, um Hybridsaatgut herzustellen.
Bei vielen kultivierten Pflanzen liegt bei der Etablierung einer Hybridsaatzucht die Schwierigkeit in der Tatsache, daß sie hermaphroditisch sind, also Blüten mit männlichem und weiblichem Geschlecht aufweisen. Bei den Getreiden, deren Blüten winzig sind, verursacht das erhebliche praktische Probleme bei Kreuzungen. Angestrebt wird daher die Etablierung von Linien, die männlich steril sind, sowie Linien, in denen diese Sterilität wieder aufgehoben ist (Restorer). Letztere dienen dann auch der Vermehrung der männlich sterilen Inzuchtlinien.
Heute können solche Linien gentechnologisch hergestellt werden.
Hier soll ein Szenario angedeutet werden, das von der belgischen Firma PGS unter Zuhilfenahme gentechnologischer Methoden entwickelt wurde (2). Die Tapetum-spezifische Ausprägung einer bakteriellen RNAse (Barnase) in den Antheren transgener Tabakpflanzen verhindert die Reifung der Pollen und führt somit zur männlichen Sterilität. Die genetische Erhaltung dieser Linie ist nur dann möglich wenn die Sterilität rückgängig gemacht werden kann. Die gentechnologische Einführung und tapetal-spezifische Ausprägung eines Inhibitors der RNAse (Barstar) hebt die Sterilität auf (Restorer) und die Pflanze kann generativ vermehrt werden. Dieses System wurde von PGS inzwischen auch für Raps ausgetestet.
Andere Systeme männlicher Sterilität nutzen die Möglichkeiten des Sekundärmetabolismus der Pflanzen, indem z.B. die Bildung von Flavonolen in den Tapetalzellen transgener Pflanze durch anti-sense Expression des Chalkonsynthase-Gens verhindert wird (3). In Abb.5 ist dies verallgemeinernd durch die Ausprägung eines Sterilitätsgens dargestellt, das gentechnologisch eingeführt wurde und die Entwicklung von fertilem Pollen verhindert. Linie A kann zusätzlich mit dem Gen Tnp A transformiert werden, so daß dessen Genprodukt die Ausprägung des Sterilitätsgens blockiert, was zur Restaurierung der männlichen Fertilität führt. Linie B kann dann geselbstet und zu einer Inzuchtlinie entwickelt werden. Die segregierenden männlich sterilen Linien können für die entscheidende Kreuzung mit einer anderen analog hergestellten Inzuchtlinie zur Erstellung des Hybridsaatgutes verwendet werden. Dieses System wird gegenwärtig von der holländischen Firma De Laat den Züchtern zur Nutzung angeboten. Da sich dieses Angebot jedoch auf die Genkonstrukte und die Primärtransformanten und nicht notwendigerweise auf "fertige " Kultivare bezieht, wartet auf den Züchter noch viel Arbeit.
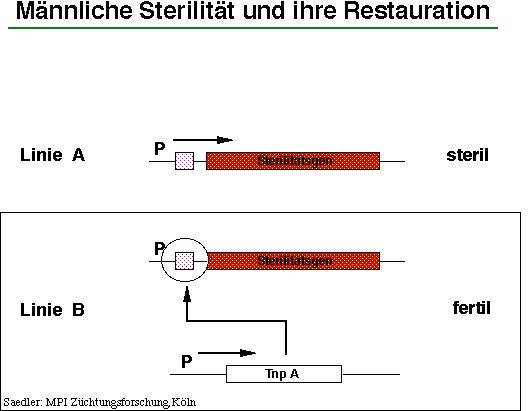
Männliche Sterilität und ihre Retauration
Wie einleitend bereits erwähnt sind die Ernteverluste in Entwicklungsländern durch Pflanzenkrankheiten besonders hoch. Verluste durch virale Erkrankungen sind insbesondere in Afrika erheblich. Eine chemotherapeutische Behandlung der Pflanzen ist derzeit nicht möglich. Schutzmaßnahmen beschränken sich auf die prophylaktische Bekämpfung der überträger mittels Insektizide. Hierfür fehlen den Entwicklungsländern aber oft die Mittel.
Der Anbau von virus-resistenten Pflanzen wäre wünschenswert und könnte Ernteverluste in den Entwicklungsländern reduzieren und somit zur Erhöhung der zur Verfügung stehenden Nahrungsmittel beitragen.
Die gentechnologische Herstellung von virus-resistenten Pflanzen wird weiter unten beschrieben. Diese erfordert aber auf jeden Fall molekularbiologische Hochtechnologie.
Der immer wiederkehrenden Behauptung, High-Tech sei für Entwicklungsländer unbezahlbar, könnte folgendermaßen begegnet werden:
Die Erstellung der transgenen Unikate könnte z.B.in der Bundesrepublik vorgenommen, die Einkreuzung in die Landsorten vor Ort vorgenommen werden. Da in diesem Szenario Märkte hiesiger Unternehmen nicht oder kaum berührt sind, ist auch auf diesem Sektor eine Kooperation zwischen Industrie und öffentlicher Hand denkbar. Die transgenen Unikate würden im Rahmen der Entwicklungshilfe den Drittweltändern überlassen. Entwicklunsghilfe bedeutet also hier: Schaffung von Arbeitsplätzen in der Bundesrepublik und (unentgeldlicher) Abgabe des Produktes an die Drittweltländer.
Dies scheint nicht nur eine neue, sondern auch eine effektive Art von Entwicklungshilfe zu sein, da neben der Lösung von Problemen vor Ort, deutsche Strukturen auf- bzw.ausgebaut, in jedem Fall aber bereits sehr frühzeitig in den Prozeß einer Entwicklungshilfe miteinbezogen werden.
Im folgenden sollen Schlüsselthemen Zentraleuropas angesprochen werden, die weniger einer Erhöhung der Nahrungsmittelproduktion als vielmehr der Nahrungsqualität und insbesondere ökologischen Problemen der Landwirtschaft gewidmet sind.
Der Anbau von genetisch einheitlichen Pflanzen, wie er in der intensiven Landwirtschaft heute üblich ist, zieht eine Vielzahl von Umweltproblemen nach sich. Erwähnt wurde der Verlust der Kompetitivität der Kultivare, was zur Folge hat, daß diese vor ihren "Mitbewerbern" auf den Agrarflächen geschützt werden müssen. Dies geschieht durch die Ausbringung von Herbiziden, also Wirkstoffen, die das Wachstum unerwünschter Pflanzen auf der Nutzungsfläche verhindern sollen. Die heute verwendeten Herbizide wirken spezifisch auf "Unkräuter" und beeinflussen nicht den zu schützenden Kultivar. Die hohen Aufwandmengen und die lange Bodenpersistenz vieler derartiger Herbizide kann zu einem Eindringen in das Grundwasser führen. Dies kann nicht länger hingenommen werden. Infolgedessen bemüht sich die Industrie denn auch neuartige, kurzlebigere Substanzen zu entwickeln, die in geringeren Aufwandmengen ausgebracht werden können. Leider fehlt diesen "neuen" Herbiziden jedoch oftmals die spezifische Wirkung, d.h. sie inaktivieren alle Pflanzen und werden daher als Totalherbizide bezeichnet. Sie sind somit nur vor Auflauf der Saat, also zeitlich sehr eingeschränkt verwendbar.
Eine umfassendere Nutzung derartiger Substanzen erscheint ökologisch sinnvoll, kann aber nur angegangen werden, wenn es gelingt, den Kultivar vor der Wirkung dieser Substanzen zu schützen.
Bevor wir jedoch hierauf eingehen, seien einige anderen Probleme intensiver Landwirtschaft kurz angesprochen. Der Anbau von Kurzstrohsorten, aber auch anderer Hochertragssorten ist nur bei ausreichender Applikation von Dünger besonders produktiv. Hierin liegt aber wiederum ein anderes ökologisches Problem, da zunehmend z.B. mehr Nitrate in das Grundwasser gelangen. Darüberhinaus stellt der Anbau von genetisch einheitlichem Material eine ideale Grundlage für die explosionsartige Ausbreitung von Pflanzenkrankheiten dar, die nur durch erhöhten Einsatz von Pflanzenschutzmitteln kontrollierbar sind. Dies ist eine weitere Quelle ökologischer Belastungen.
Die klassischen, in der Öffentlichkeit vieldiskutierten Szenarien gentechnisch veränderter Pflanzen beziehen sich auf Herbizid-Toleranz und erhitzen die Gemüter.
Die Bodenpersistenz der Herbizide der neuen Generation liegt deutlich unter der der jetzt verwendeten. Infolge ihrer mangelnden Wirtsspezifität müssen die Kultivare jedoch vor deren Wirkung geschützt werden. Dies ist mit Hilfe gentechnologischer Methoden möglich.
Viele der neuen Wirkstoffe,die von Mikroorganismen produziert werden, greifen z.B. in die Aminosäurebiosynthese ein, so daß "alle" Pflanzen empfindlich gegenüber diesen Wirkstoffen sind. Allerdings schützen sich die Bakterien vor ihren Wirkstoffen mit Hilfe eines Resistenz-Gens. Eine derartige Toleranz kann auf mehreren Mechanismen basieren. Einmal wird das "sensitive" Enzym überproduziert und somit eine Toleranz gegenüber einer bestimmten Dosis des Wirkstoffes erreicht, zum anderen kann ein mutiertes Enzym vorliegen, das unempfindlich gegenüber der Wirkungsweise der Substanz ist und zum dritten kann eine neue Enzymaktivität in dem Bakterium vorliegen, die den Wirkstoff inaktiviert. Gene aus solchen toleranten Bakterien wurden isoliert, in ihren Expressionssignalen verändert und in Pflanzen eingebracht. Das Ergebnis war, daß diese jetzt tolerant gegenüber dem Herbizid wurden (4).
Der Einsatz derartiger transgener Pflanzen könnte zu einer Reduktion der Aufwandmengen an Herbiziden in der Landwirtschaft führen, da nunmehr äußerst effektive, aber relativ kurzlebige Wirkstoffe nur nach entsprechender Indikation eingesetzt werden müßten. Die Belastung der Ökosysteme sänke entsprechend.
Der Einsatz von Pestiziden, hier insbesondere Insektizide hat ein bedrohliches Ausmaß erreicht, so daß auch hier nach neuen Wegen zu suchen ist. Seit vielen Jahren werden biologische Präparate zur Bekämpfung von Insektenfraß eingesetzt. Unterschiedliche Bazillus thuringiensis (Bt) Arten produzieren äußerst spezifische Toxine, die sehr spezifisch auf bestimmte Insektengruppen wirken. Aus diesem Grund wurden die entsprechenden Mikroorganismen auf besonders gefährdete Kultursorten, etwa Baumwolle ausgebracht, um eine möglichen Schädigung zu verhindern. Viel eleganter und auch sicherer wäre eine Verlagerung dieser Schutzmaßnahme direkt in die Pflanze. Toxin-kodierende Gene wurden aus verschiedenen Bac.thuringiensis Arten isoliert und in ihren Expressionssignalen so verändert, daß eine Ausprägung auch in Pflanzen stattfinden konnte. Transgene Bt-Pflanzen sind, wie viele Versuche gezeigt haben, resistent gegenüber Fraß durch bestimmte Insekten.
In großflächigen Freilandversuchen, insbesondere in den USA, konnte so bis zu einem Drittel an Pestiziden eingespaart werden (5).
Dies scheint ein Schritt in die richtige Richtung zu sein.
Leider ist in Deutschland die Bereitschaft derzeit noch nicht vorhanden, diese positiven Seiten, der neuen Technologien zu nutzen. Hier gilt es noch viele Vorurteile, die größtenteils auf Unkenntniss basieren, abzubauen.
Große Teile der Ernte sind in jedem Jahr durch virale Erkrankungen gefährdet. Da hier keine chemischen Wirkstoffe existieren, die eine Therapie ermöglichen, können nur die überträger der Krankheiten, oftmals Insekten, durch Pestizide bekämpft werden. Dieser ungezielte, oft auch prophylaktische Einsatz von hochwirksamen Substanzen belastet die Umwelt und sollte soweit als irgend möglich eingeschränkt werden.
Bislang hatte der Züchter lediglich die Möglichkeit Virus-Resistenz-Gene, soweit sie in verwandten und kreuzbaren Arten bekannt waren, in seine Kultursorten einzukreuzen. Dies ist jedoch ein zeitaufwendiges Verfahren und bringt oftmals nur geringen Erfolg, da nur wenige Virus-Resistenzen monogen vorliegen.
Hier eröffnet die Gentechnologie neue Möglichkeiten.
Eine Vielfalt von Virus-resistenten Kulturpflanzen wurde gentechnisch erzeugt (6) und in vielen Freilandversuchen getestet.
Hier sollen keine Einzelheiten aller Möglichkeiten beschrieben werden, die die Etablierung einer Virus-Resistenz in einem Kultivar ermöglichen. Vielen Strategien gemeinsam ist, daß Segmente des viralen Genbestandes in das Pflanzengenom integriert und ausgeprägt werden. Dies verleiht der Pflanze, auf ganz unterschiedlichen Ebenen der Vermehrung und Verbreitung des Virus, einen Schutz. Dies sei am Beispiel der Blattrollvirose bei Kartoffeln dargestellt, da entsprechende PLRV-resistente Pflanzen 1994 am Max-Planck-Institut für Züchtungsforschung im Freiland getestet werden (7).
PLRV kodiert ein Protein, das für seine Ausbreitung in der Pflanze durch das Phloem benötigt wird. Hierbei bindet eine Domäne des Proteins an die Nukleinsäure des Virus und eine andere Domäne dient der Polymerisation der Proteine untereinander auf der Virus Nukleinsäure. Dies ermöglicht die Ausbildung von gestreckten Strukturen und somit einen Transport von Zelle zu Zelle durch die Plasmodesmata. Das virale Gen wurde isoliert und so verändert, daß das von ihm kodierte Protein bei Co-Polymerization auf der viralen Nukleinsäure keine gestreckten Strukturen mehr bildet und daher der Transport von Zelle zu Zelle unterbleibt. Kartoffeln, die dieses Genkonstrukt enthalten, sind im Gewächshaus resistent gegenüber dem Blattroll-Virus (PLRV).
Für eine Nutzung dieser Resistenz in der Praxis bedarf es aber natürlich noch der Erprobung im Freiland.
Der Anbau Virus-resistenter transgener Pflanzen eröffnet die Möglichkeit, virale Erkrankungen, die wie im Fall des Blattroll Virus bis zu 80%-ige Ernteverluste bedingen, auch wirksam bekämpfen zu können. Dies könnte wiederum helfen Insektizide einzusparen, was wiederum als Beitrag zur Entspannung der Umweltproblematik zu sehen ist.
Eine weiteres großes Problem der Landwirtschaft industrialisierter Länder stellen Pilzkrankheiten dar. Insbesondere durch den Anbau von Monokulturen besteht die Notwendigkeit, Fungizide zur Bekämpfung auftretender Pflanzenkrankheiten einzusetzen. Dies hat zur Folge, daß zunehmend Umweltschädigungen beobachtet werden. Die Züchter stellen sich zunehmend darauf ein, Krankheitsresistenzgene aus verwandten Wildsorten in die Kultivare einzukreuzen.
Der Anbau krankheitsresistenter Sorten würde eine erhebliche Einsparung sonst notwendiger Pflanzenschutzmittel nach sich ziehen.
Die Erstellung biologisch geschützter Linien ist, wie bereits vorhin erwähnt wurde, eine sehr zeitraubende Arbeit, die jedoch mit Hilfe von High-Tech wie etwa der RFLP-Methoden zielgerichteter und somit auch schneller durchgeführt werden könnte. Dies setzt allerdings voraus, daß der Züchter, die entsprechenden High-Tech Methoden auch nutzen kann.
Die Suche des Züchters nach geeigneten Nachkommen einer Kreuzung erfolgt klassischerweise durch die Auswahl auf phänotypischer Ebene, also bei der Ausprägung eines Merkmals. Auch hier hat die Molekularbiologie eine methodische Vielfalt an Alternativen entwickelt, die es ermöglicht, eine gesuchte Rekombinante auf genetischer Ebene bereits in einer 2.Filialgeneration zu ermitteln. Sogenannte RFLP-(Restriction-Fragment-Length-Polymorphism) Analysen leisten dies und sind heute Bestandteil des Methodenrepertoires moderner international erfolgreicher Pflanzenzüchter. Diese Methode ist dann leistungsfähig, wenn eine hinreichend große Zahl von Pflanzen analysiert werden kann. Entsprechendes Know-how und die notwendige Kapazität fehlt aber vielen deutschen Pflanzenzüchtern. Einige moderne Methoden, wie z.B. die AFLP-Technik, sind sogar noch weitaus leistungsfähiger als die RFLP-Technik.
Für viele Kultivare mögen entsprechende resistente Wildarten nicht zur Verfügung stehen; dann könnte ein Resistenzgen aus irgendeiner Pflanze gentechnologisch isoliert werden und stünde somit zur übertragung auf recht unterschiedliche Pflanzenarten bereit, die sich dadurch alle besser gegenüber pilzlichen Erkrankungen schützen ließen.
Die Molekularbiologie hat Methoden entwickelt, die es erlauben, Gene aus Mikroorganismen, Tieren oder Pflanzen zu isolieren, sie in-vitro zu verändern und z.B. wieder in Kulturpflanzen einzubringen. Dies führt zu einer Bereicherung des Genbestandes des Kultivars, da er jetzt Merkmale auch aus einer anderen Spezies besitzen kann.
Leider ist derzeit die übertragung eines Gens in das Genom einer Pflanze noch nicht zielgerichtet, d.h. der Integrationsort im Pflanzengenom ist zufällig. Hierdurch bedingt kann einerseits die Ausprägung des übertragenen Gens nicht optimal oder sogar defekt sein, was etwa durch DNA-Methylierung des übertragenen Gens hervorgerufen werden kann. In jedem Fall wird der Züchter nur die transgene Pflanze auswählen, in der das Transgen optimal ausgeprägt wird und in der keine weiteren Veränderungen festzustellen sind.
Diese Technologie ist kapitalintensiv und erfordert ein Know-how, das nur bei wenigen großen deutschen Pflanzenzüchtern anzutreffen ist.
Gegenwärtig stellt die Isolierung solcher Krankheitseresistenzgene noch ein erhebliches Problem dar, denn in vielen Fällen scheint Resistenz gegen pilzliche Erreger multifaktoriell bedingt zu sein.
Im Sommer 1990 wurde in der Bundesrepublik Deutschland mit einer transgenen Petunie der erste Freilandversuch durchgeführt (8). Obwohl zu dieser Zeit keine gesetzliche Regelung existierte, war dieser Versuch bereits 1989 im Umweltausschuß des Rats der Stadt Köln öffentlich zur Diskussion gestellt worden. Es wurde bald klar, daß in Deutschland einige politische Gruppen aber auch Umweltverbände gegen derartige Neuerungen mobil machten. Ideologische Voreingenommenheit hatte die Sicht für die Fakten verstellt.
Bereits 1987 war durch übertragung eines Maisgens in eine geeignete Petunienmutante ein Biosyntheseweg eröffnet worden, der zu einem für diese Pflanze neuen Pigment führte (9). Die nunmehr lachsrot blühende Pflanze wurde zu einem wissenschaftlichen Modellsystem, da beim Anbau im Freiland neue Befunde erhoben werden konnten, die weltweite Forschungsaktivitäten initiierten.
Etwa 30 000 lachsrot blühende Petunien wurden im Sommer 1990 im Freiland angebaut. Mehr als 92% der ersten Blüten der Pflanzen zeigten die erwartete lachsrote Pigmentierung. Im Verlauf des Sommers fand jedoch eine bis dahin nicht gekannte Umprogrammierung im Erscheinungsbild der Blüten statt. Nur 30% der späteren Blüten zeigten die lachsrote Farbe. Die molekulare Analyse ergab, daß das übertragene Maisgen in mehr als 60% der Pflanzen nach starker Veränderung einiger Umweltfaktoren (langanhaltende klare, heiße Sommertage mit einem hohen UV-Anteil) einen hohen Methylierungsgrad aufwies, der, wie spätere Experimente im Labor ergab, zur Abschaltung des Maisgens führte (8).
Neben der Beobachtung eines für Petunia neuen Pigmentes und der Umwelt-induzierten DNA-Methylierung konnte ein weiterer Befund erhoben werden. Die transgenen Pflanzen waren erheblich resistenter gegenüber einer Vielzahl an Krankheitserregern als die weißblühende Ausgangslinie (10). Diese akkumulierte, bedingt durch Mutationen in zwei unterschiedlichen Genen, Dihydrokaempferol, das durch die petunieneigene Dihydroflavanol-Reduktase nicht umgesetzt werden kann. Das entsprechende Maisenzym hat keine derartige Substratspezifität und wandelt daher die obige Substanz in Leukopelargonidin um. Die nachgeschalteten Petunienenzyme können dieses Substrat zu einem gewissen Anteil in Folgeprodukte und schlußendlich in das für Petunia neue Pigment Pelargonidin umwandeln. Dies führt zu der lachsroten Blütenfarbe. Infolge der konstitutiven Ausprägung des Maisgens und des reduzierten Abflußes des gebildeten Leukopelargonidins in den Blättern könnte dort diese Substanz spontan polymerisieren und somit Gerbstoffe, wie etwa Proanthozyanidine bilden. Diese Substanzen inaktivieren und präzipitieren Proteine und sind somit auch für Pflanzen möglicherweise toxisch. Ein möglicher Schutz für die Pflanze wäre ein Transport in die Vakuole. Sollte allerdings ein Krankheitserreger die Zelle befallen, könnte der Inhalt der Vakuole nicht nur die Pflanzen-, sondern auch die Pilzzelle abtöten oder sie zumindest in ihrem Wachstum hemmen. Pflanzen mit diesen Inhaltsstoffen sollten demnach resistent gegenüber vielen Pathogenen sein. Diese Vorstellungen sind nicht bewiesen, werden jedoch durch die Befunde mit den transgenen Petunien, aber auch durch alte Beobachtungen von Züchtern nahegelegt (11). Die transgenen Petunien zeigen eine quantitative Resistenz gegenüber vielen pilzlichen Krankheitserregern (Abb.6).
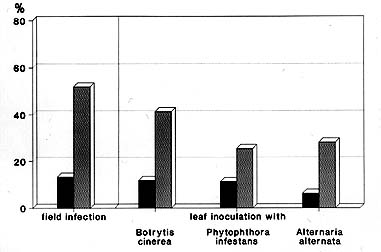
Breitspektrum-Resistenz
Infolge der gerbstoffartigen Wirkung von Proanthozyanidinen ist deren Konsum für monogastrische Tiere nicht angezeigt. Wildpflanzen wehren sich gegen Pathogenbefall häufig durch derartige Sekundärstoffe.
Da inzwischen die notwendigen Gene für eine Manipulation dieses Biosyntheseweges bekannt und isoliert worden sind, kann mit den Methoden der Gentechnik die Biosynthese dieser Stoffe in anderen Pflanzen aufgebaut und gesteuert werden, so daß jetzt erstmals die Synthese einer Breitspektrum-Resistenz möglich ist.
Biosynthesewege anderer Sekundärstoffe müssen weiter erforscht und können dann möglicherweise analog genutzt werden. Auch hier ist ein Zusammenspiel zwischen Forschung und Anwendern angezeigt, möglicherweise ist für die Nutzung der Erkenntnisse eine separate Institution für die Anpassung der Ergebnisse an die Praxis notwendig.
Im letzten Beispiel geht es um ein futuristisch anmutendes Projekt.
In Abb.7 ist vereinfachend die Beziehung zwischen der Photosynthese und der Atmung der Lebewesen dargestellt. Ohne menschlichen Einfluß befinden sich diese beiden Prozesse im Gleichgewicht. 35 Milliarden Tonnen Kohlendioxyd, die bei der Atmung entstehen, werden durch die Photosynthese wieder gebunden. Die Verbrennung fossiler Ressourcen sowie das Abholzen insbesondere der Regenwälder führt zu einer empfindlichen Störung dieses Kreislaufes, so daß nun ein überschuß an Kohlendioxyd entsteht, der möglicherweise auch zu einer Aufheizung der Atmosphäre beitragen kann. Nachwachsende Rohstoffe könnten zur Entspannung dieser Problematik beitragen. Hier könnten u.a. Rapssorten eingesetzt werden, deren ölgehalt und/oder Fettsäurezusammensetzung gentechnologisch zu verändern wäre.
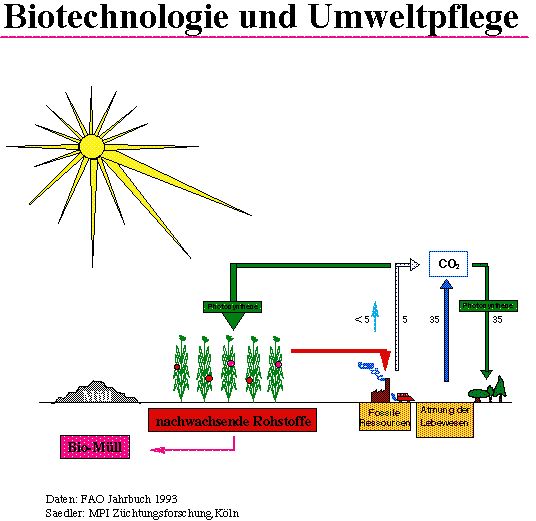
Biotechnologie und Umweltpflege
Aber es kämen auch andere transgene Pflanzen in Frage, deren Inhaltsstoffe zu neuen, die Umwelt weniger belastenden Produkten (Bioplastik) führen könnten. Bakterien der Art Alcaligenes eutrophus produzieren Polyhydroxybuttersäure, ein thermisch verformbares Material, das den großen Vorteil hat, biologisch abbaubar zu sein und daher kurz als Bioplastik bezeichnet wird. Bislang ist seine Produktion in Bakterien zu teuer, um mit chemisch produzierten Plastikmaterialien konkurrieren zu können. Gelänge eine Produktion dieses Materials in einer Kulturpflanze, dann könnte der große Vorteil der biologischen Abbaubarkeit voll genutzt werden und somit eines unserer dringlichen Umweltprobleme (Müll) lösen helfen. Für die Biosynthese von Polyhydroxybuttersäure sind drei Enzyme notwendig, deren Gene von der Arbeitsgruppe um Chris Somerville aus Bakterien isoliert und nach entsprechender Veränderung in Arabidopsis thaliana eingebracht wurden (12). Die Keimlinge akkumulieren das Bioplastikmaterial in kleinen Kügelchen, aber leider im Zellkern, so daß der Keimling eingeht. Dieses Problem gilt es zu lösen, bevor Aussicht auf eine Anwendung besteht.
Dennoch zeigen diese Versuche, welche Potenz in dieser neuen Technologie liegt und welche Hilfe sie bei der Lösung vieler Probleme bieten könnte.
Woran liegt es dann, daß die Vorteile dieser Technologie in der Bundesrepublik Deutschland nicht oder nur zögernd genutzt werden? Stellt die Nutzung gentechnologisch hergestellter Pflanzensorten ein neues Risiko dar?
Von den mehr als 1300 Freilandversuchen mit transgenen Pflanzen, die bis 1993 weltweit durchgeführt wurden, fanden nur wenige in Deutschland statt (Abb.8).

Freilandversuch mit trangenen Pflanzen
Die Gründe hierfür sind sicherlich vielfältig; neben rein bürokratischen Hürden spielt sicherlich die mangelnde öffentliche Akzeptanz dieser neuen Technologie eine erhebliche Rolle. Im wesentlichen scheint es dabei um Vorurteile zu gehen, die auf der Annahme basieren, daß transgene Pflanzen sich unkontrolliert ausbreiten und somit das "biologische Gleichgewicht" erheblich verschieben könnten. Beispielhaft hierfür wird dann auch immer die unkontrollierte Ausbreitung von Wildpflanzen angeführt, wenn diese in neue Biotope verbracht werden.
Wildpflanzen stellen aber natürlich keine guten Modellsysteme für diese Frage dar, denn im Gegensatz zu den kultivierten Pflanzen besitzen sie alle Eigenschaften, die eine optimale Ausbreitung ermöglichen.
Bei der Domestizierung der Pflanzen (siehe Einleitung) wurden in der Regel Pflanzen ausgewählt, die in ihrem Verbreitungspotential drastisch eingeschränkt sind. Beim Mais z.B. sind die Körner so fest am Kolben anhaftend, daß sie nur noch mit Gewalt abgetrennt werden können. Daher kann Mais nur noch durch Menschenhand verbreitet werden. Entsprechendes gilt für viele andere Kulturpflanzen.
Entstanden sind unsere Kulturpflanzen auf den verschiedenen Kontinenten. Sie werden heute jedoch weltweit angebaut, oftmals ohne daß Verwilderung zu beobachten ist. In Abb.9 ist die Verwilderungstendenz wichtiger Kulturpflanzen Mitteleuropas aufgeführt (13). Klar ersichtlich ist, daß keine nennenswerten Verwilderungen bislang beobachtet wurden, obwohl diese Pflanzen seit Jahrhunderten im Anbau sind, wie etwa die "Exoten" Mais oder Kartoffel, die beide aus Latein-Amerika stammen. Selbst mit Raps, bei dem eine Verwilderungstendenz möglich erscheint, konnte in 3-jährigen Anbauversuchen mit transgenem Material keine Invasion unterschiedlicher Habitate beobachtet werden (14).

Verwilderung von Kuturpflanzen
Neben der Ausbreitung der Kultursorte per se könnten sich Gene auch durch Pollenflug auf einheimische Arten ausbreiten, falls diese miteinander kreuzbar wären. Dies ist jedoch bei vielen der aufgeführten Kulturpflanzen nicht der Fall. Lediglich bei den einheimischen Kultivaren Raps und Zuckerrübe bestünde ein diesbezügliches Ausbreitungspotential, da hier nahe verwandte Pflanzen ebenfalls einheimisch sind.
Gentechnologische Veränderungen werden vornehmlich an Kulturpflanzen mit geringem Verbreitungspotential vorgenommen.
So kann etwa der von Menschenhand geschaffene Mais, der in der freien Natur nicht anzutreffen ist, als Sicherheitssystem par excellence angesehen werden. Gentechnologisch veränderter Mais, unabhängig von dem eingebrachten Gen, stellt somit keinerlei erhöhtes Risiko für die Umwelt dar. Ähnliches gilt für die Kartoffel, die Tomate und andere. Im Falle des Hafers oder des Raps etwa könnte aber die Funktion der transferierten Gene bei einer Risikoabwägung eine Rolle spielen.
Von daher scheint für eine landwirtschaftliche Nutzung gentechnologisch veränderter Pflanzen die Aufstellung einer Positiv-Liste der Pflanzen, deren Ausbreitungspotential unbedenklich ist, angezeigt.
Wenn eine europäische Kompetetion mit außer europäischen Landwirtschaften erfolgreich sein soll, dann sind nicht nur nationales Recht, sondern auch EU Richtlinien entsprechend zu ändern.
Dieser Vortrag erschien in:
Wissenschaft in der globalen Herausforderung;17.-20 Setember 1994;hrsg. von Hubert Markl..., Mit Betr. von E.Arzt,Stuttgart:Wiss. Verlagsgesellschaft Stuttgart 1995; S.Hirzel-; S.235-249 Edition UNIVERSITAS